Zhang Yuxun, Bharathi Sivakama S, Rardin Matthew J, Uppala Radha, Verdin Eric, Gibson Bradford W, Goetzman Eric S
Department of Pediatrics, University of Pittsburgh, Children's Hospital of Pittsburgh, Pittsburgh, Pennsylvania, United States of America.
Buck Institute for Research on Aging, Novato, California, United States of America.
PLoS One. 2015 Mar 26;10(3):e0122297. doi: 10.1371/journal.pone.0122297. eCollection 2015.
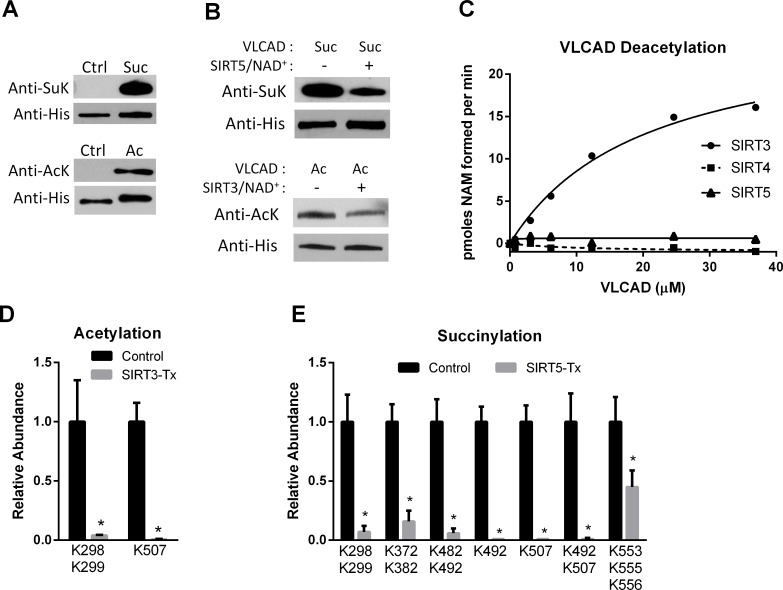
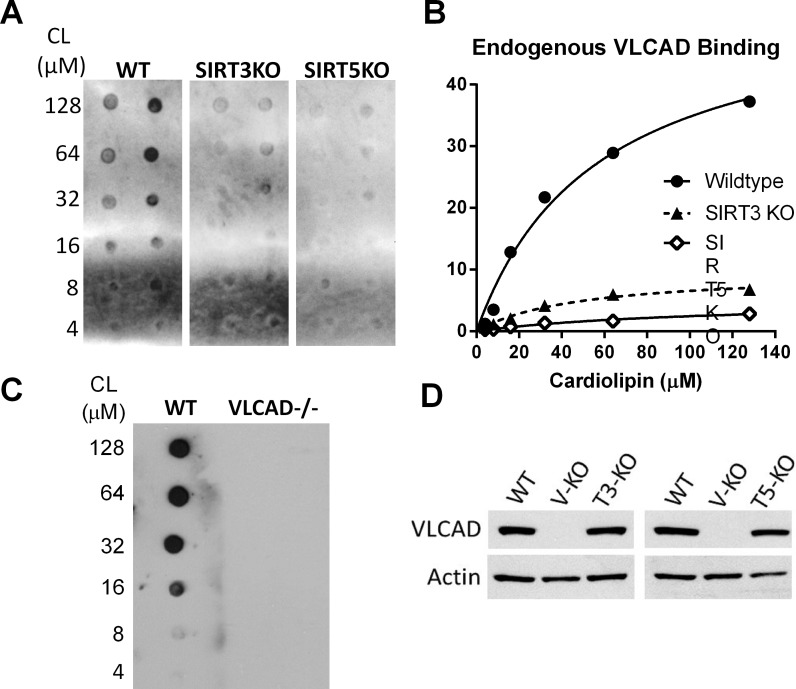
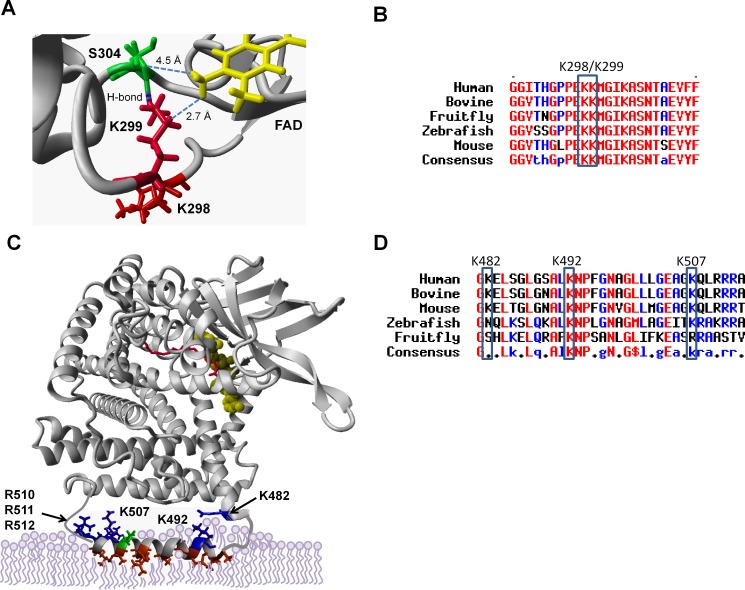
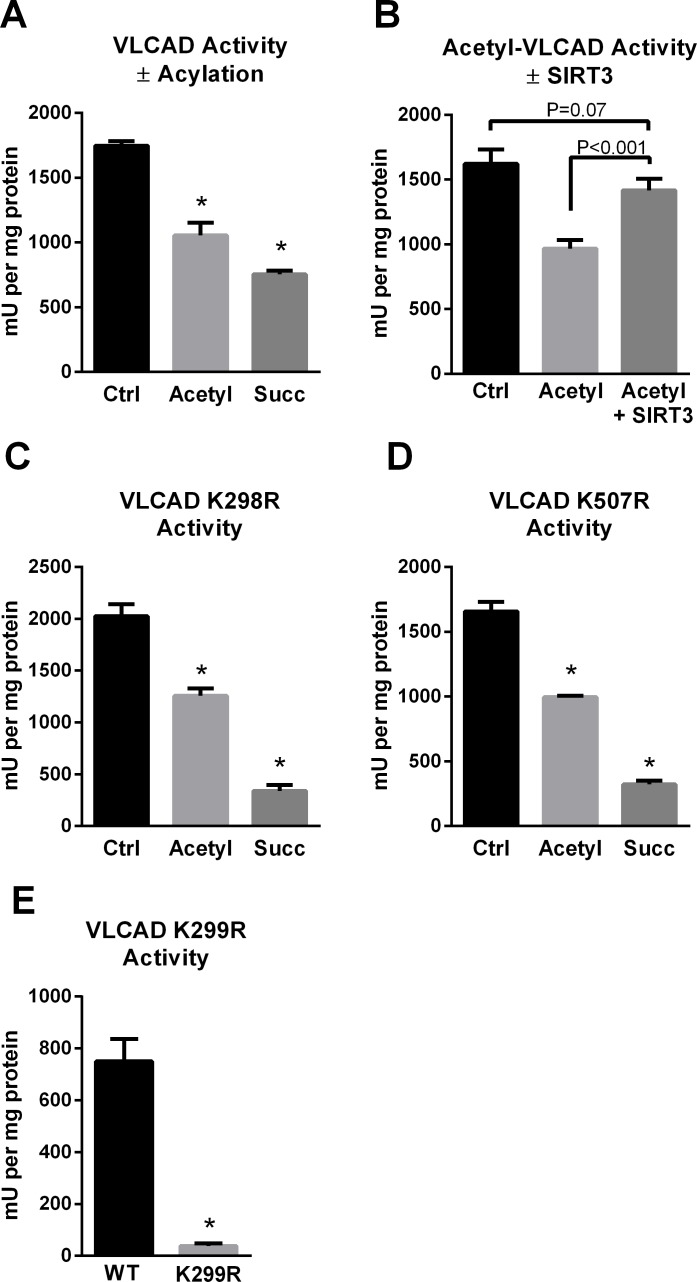
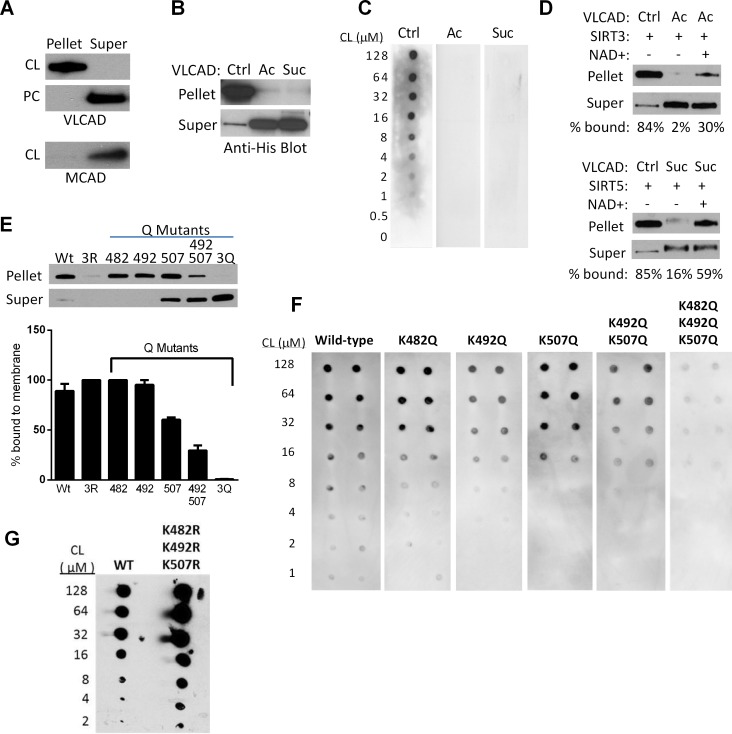
SIRT3 and SIRT5 have been shown to regulate mitochondrial fatty acid oxidation but the molecular mechanisms behind the regulation are lacking. Here, we demonstrate that SIRT3 and SIRT5 both target human very long-chain acyl-CoA dehydrogenase (VLCAD), a key fatty acid oxidation enzyme. SIRT3 deacetylates and SIRT5 desuccinylates K299 which serves to stabilize the essential FAD cofactor in the active site. Further, we show that VLCAD binds strongly to cardiolipin and isolated mitochondrial membranes via a domain near the C-terminus containing lysines K482, K492, and K507. Acetylation or succinylation of these residues eliminates binding of VLCAD to cardiolipin. SIRT3 deacetylates K507 while SIRT5 desuccinylates K482, K492, and K507. Sirtuin deacylation of recombinant VLCAD rescues membrane binding. Endogenous VLCAD from SIRT3 and SIRT5 knockout mouse liver shows reduced binding to cardiolipin. Thus, SIRT3 and SIRT5 promote fatty acid oxidation by converging upon VLCAD to promote its activity and membrane localization. Regulation of cardiolipin binding by reversible lysine acylation is a novel mechanism that is predicted to extrapolate to other metabolic proteins that localize to the inner mitochondrial membrane.
SIRT3和SIRT5已被证明可调节线粒体脂肪酸氧化,但背后的分子机制尚不清楚。在此,我们证明SIRT3和SIRT5均作用于人类极长链酰基辅酶A脱氢酶(VLCAD),这是一种关键的脂肪酸氧化酶。SIRT3使K299去乙酰化,SIRT5使K299去琥珀酰化,这有助于稳定活性位点中必需的FAD辅因子。此外,我们表明VLCAD通过靠近C端的一个包含赖氨酸K482、K492和K507的结构域与心磷脂和分离的线粒体膜紧密结合。这些残基的乙酰化或琥珀酰化消除了VLCAD与心磷脂的结合。SIRT3使K507去乙酰化,而SIRT5使K482、K492和K507去琥珀酰化。重组VLCAD的去乙酰化挽救了膜结合。来自SIRT3和SIRT5基因敲除小鼠肝脏的内源性VLCAD显示与心磷脂的结合减少。因此,SIRT3和SIRT5通过作用于VLCAD来促进其活性和膜定位,从而促进脂肪酸氧化。通过可逆的赖氨酸酰化调节心磷脂结合是一种新机制,预计可推广到定位于线粒体内膜的其他代谢蛋白。