Fuda Nicholas J, Guertin Michael J, Sharma Sumeet, Danko Charles G, Martins André L, Siepel Adam, Lis John T
Department of Molecular Biology and Genetics, Cornell University, Ithaca, New York, United States of America.
Department of Biological Statistics and Computational Biology, Cornell University, Ithaca, New York, United States of America.
PLoS Genet. 2015 Mar 27;11(3):e1005108. doi: 10.1371/journal.pgen.1005108. eCollection 2015 Mar.
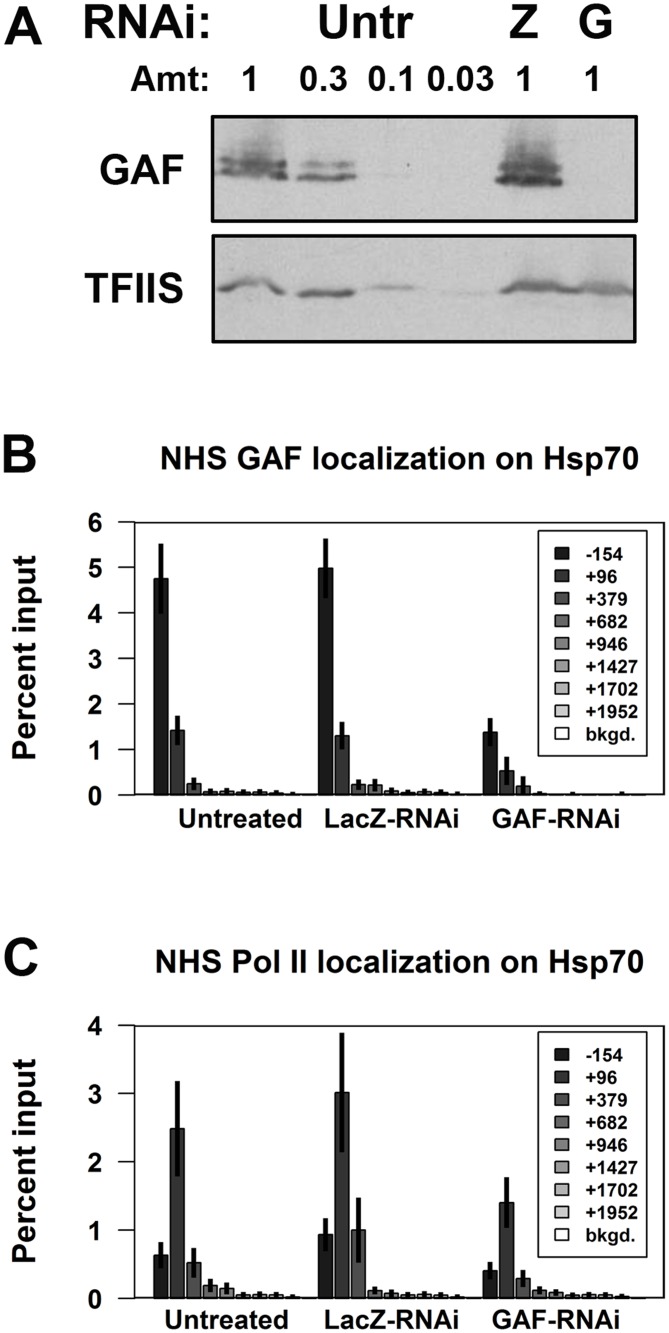
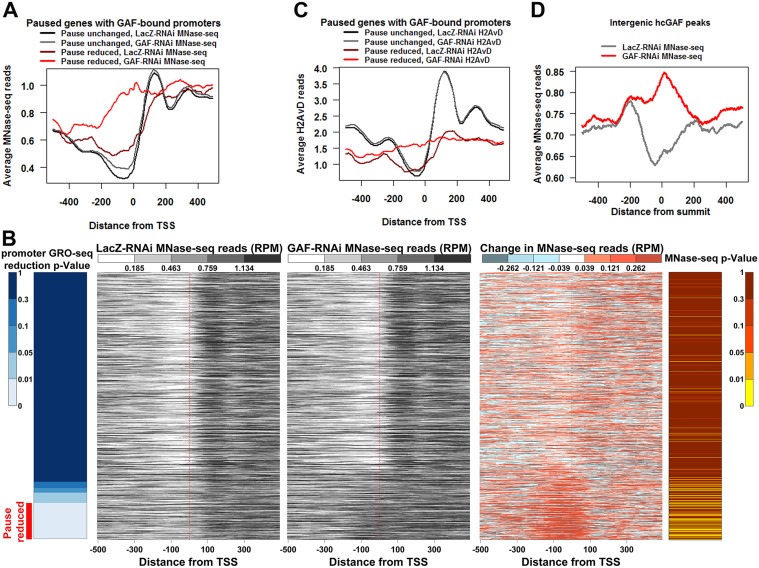
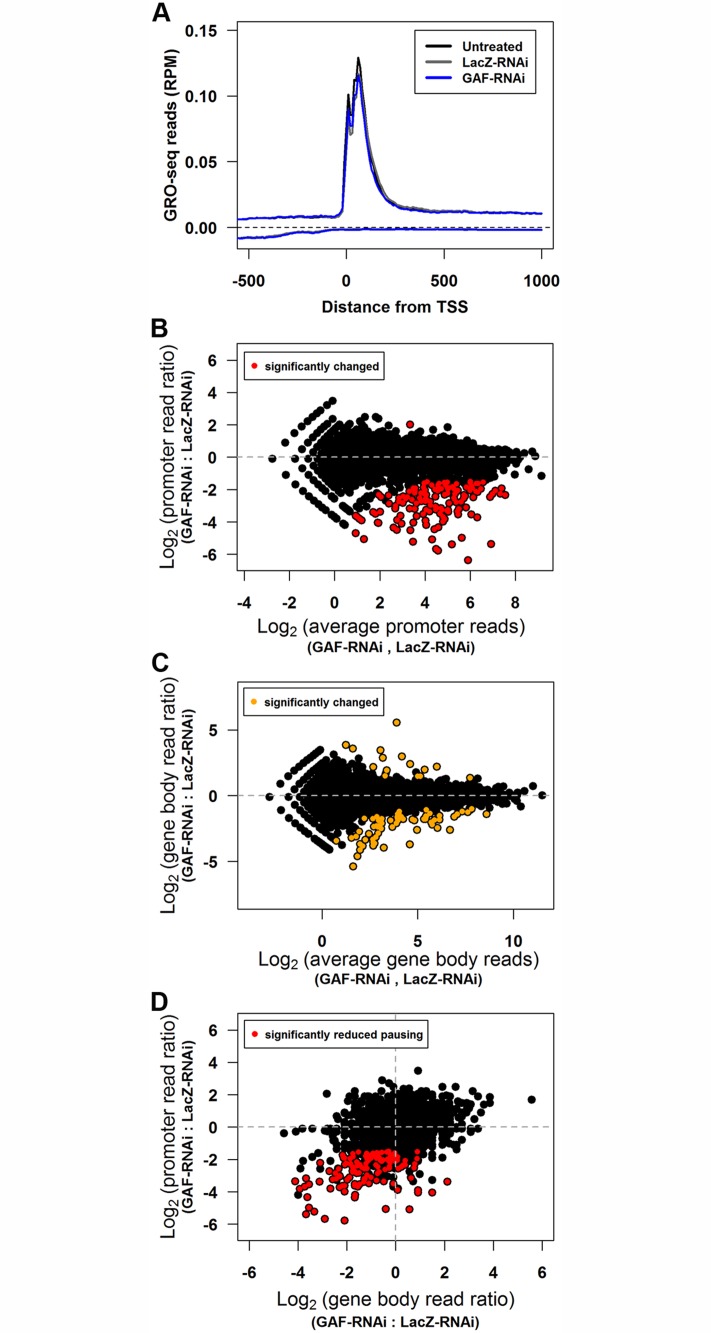
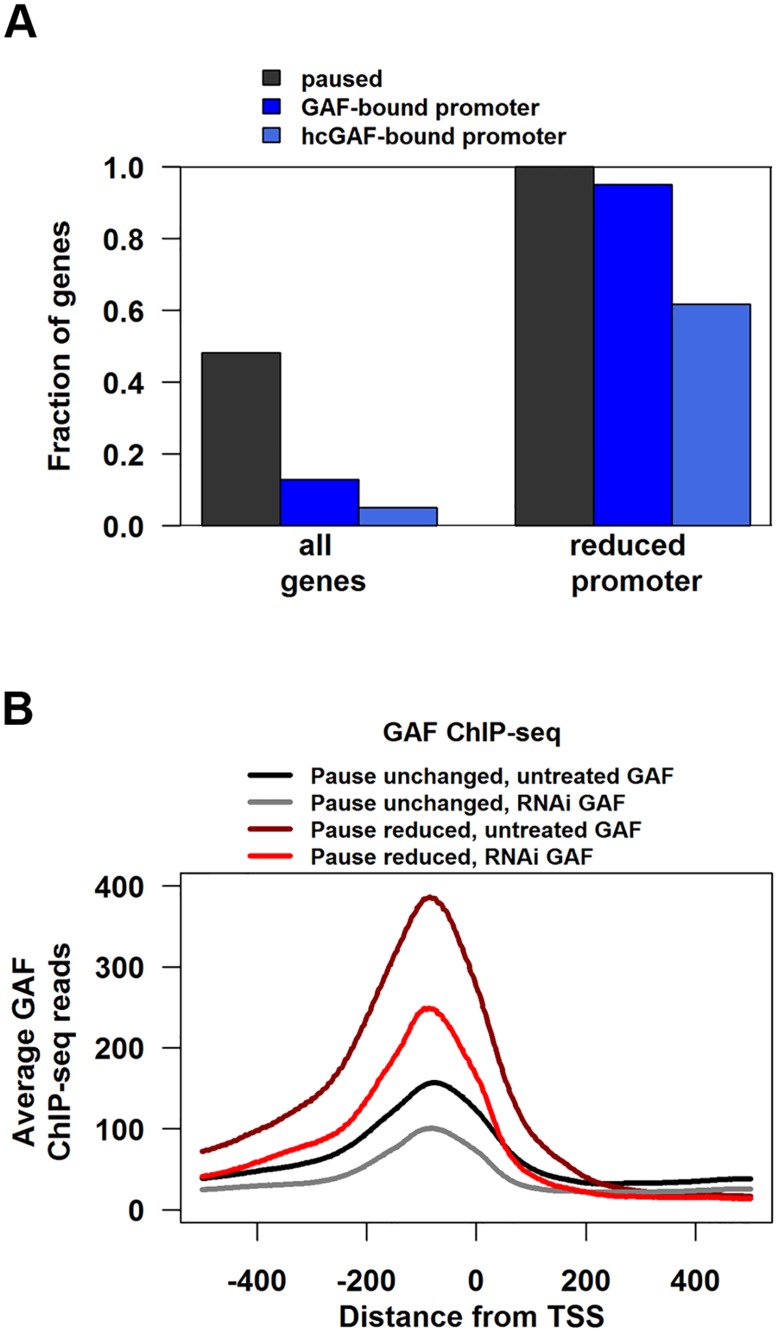
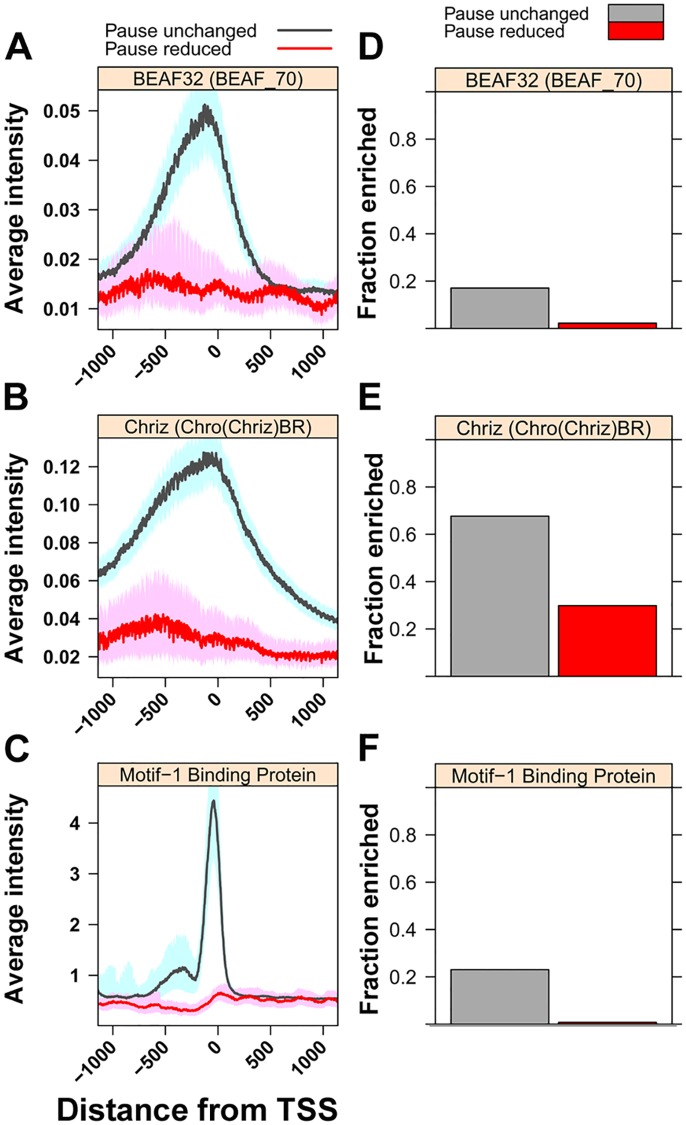
Previous studies have shown that GAGA Factor (GAF) is enriched on promoters with paused RNA Polymerase II (Pol II), but its genome-wide function and mechanism of action remain largely uncharacterized. We assayed the levels of transcriptionally-engaged polymerase using global run-on sequencing (GRO-seq) in control and GAF-RNAi Drosophila S2 cells and found promoter-proximal polymerase was significantly reduced on a large subset of paused promoters where GAF occupancy was reduced by knock down. These promoters show a dramatic increase in nucleosome occupancy upon GAF depletion. These results, in conjunction with previous studies showing that GAF directly interacts with nucleosome remodelers, strongly support a model where GAF directs nucleosome displacement at the promoter and thereby allows the entry Pol II to the promoter and pause sites. This action of GAF on nucleosomes is at least partially independent of paused Pol II because intergenic GAF binding sites with little or no Pol II also show GAF-dependent nucleosome displacement. In addition, the insulator factor BEAF, the BEAF-interacting protein Chriz, and the transcription factor M1BP are strikingly enriched on those GAF-associated genes where pausing is unaffected by knock down, suggesting insulators or the alternative promoter-associated factor M1BP protect a subset of GAF-bound paused genes from GAF knock-down effects. Thus, GAF binding at promoters can lead to the local displacement of nucleosomes, but this activity can be restricted or compensated for when insulator protein or M1BP complexes also reside at GAF bound promoters.
先前的研究表明,GAGA因子(GAF)在具有暂停的RNA聚合酶II(Pol II)的启动子上富集,但其全基因组功能和作用机制在很大程度上仍未明确。我们使用全局运行测序(GRO-seq)检测了对照和GAF-RNA干扰果蝇S2细胞中转录相关聚合酶的水平,发现敲低GAF占有率后,在大量暂停启动子的一个大子集中,启动子近端聚合酶显著减少。这些启动子在GAF缺失后核小体占有率显著增加。这些结果,结合先前表明GAF直接与核小体重塑因子相互作用的研究,有力地支持了一个模型,即GAF在启动子处指导核小体置换,从而使Pol II进入启动子和暂停位点。GAF对核小体的这种作用至少部分独立于暂停的Pol II,因为几乎没有或没有Pol II的基因间GAF结合位点也显示出GAF依赖性核小体置换。此外,绝缘子因子BEAF、与BEAF相互作用的蛋白质Chriz和转录因子M1BP在那些敲低不影响暂停的GAF相关基因上显著富集,这表明绝缘子或替代的启动子相关因子M1BP保护了一部分GAF结合的暂停基因免受GAF敲低的影响。因此,GAF在启动子处的结合可导致核小体的局部置换,但当绝缘子蛋白或M1BP复合物也存在于GAF结合的启动子时,这种活性可能会受到限制或得到补偿。