Dubrulle Julien, Jordan Benjamin M, Akhmetova Laila, Farrell Jeffrey A, Kim Seok-Hyung, Solnica-Krezel Lilianna, Schier Alexander F
Department of Molecular and Cellular Biology, Harvard University, Cambridge, United States.
Department of Mathematics, College of Science and Engineering, University of Minnesota, Minneapolis, United States.
Elife. 2015 Apr 14;4:e05042. doi: 10.7554/eLife.05042.
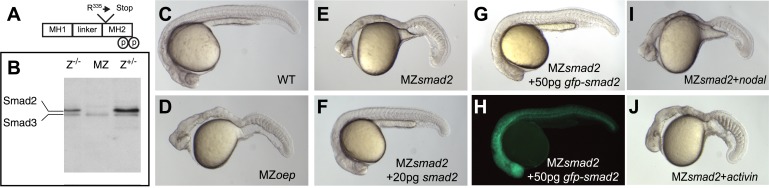
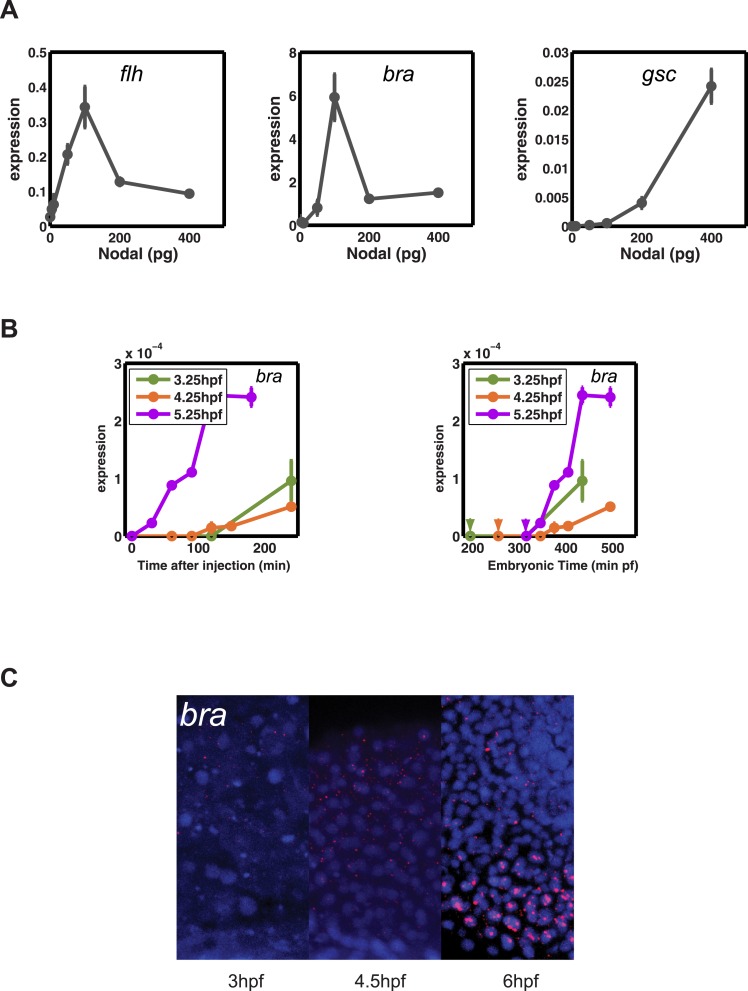
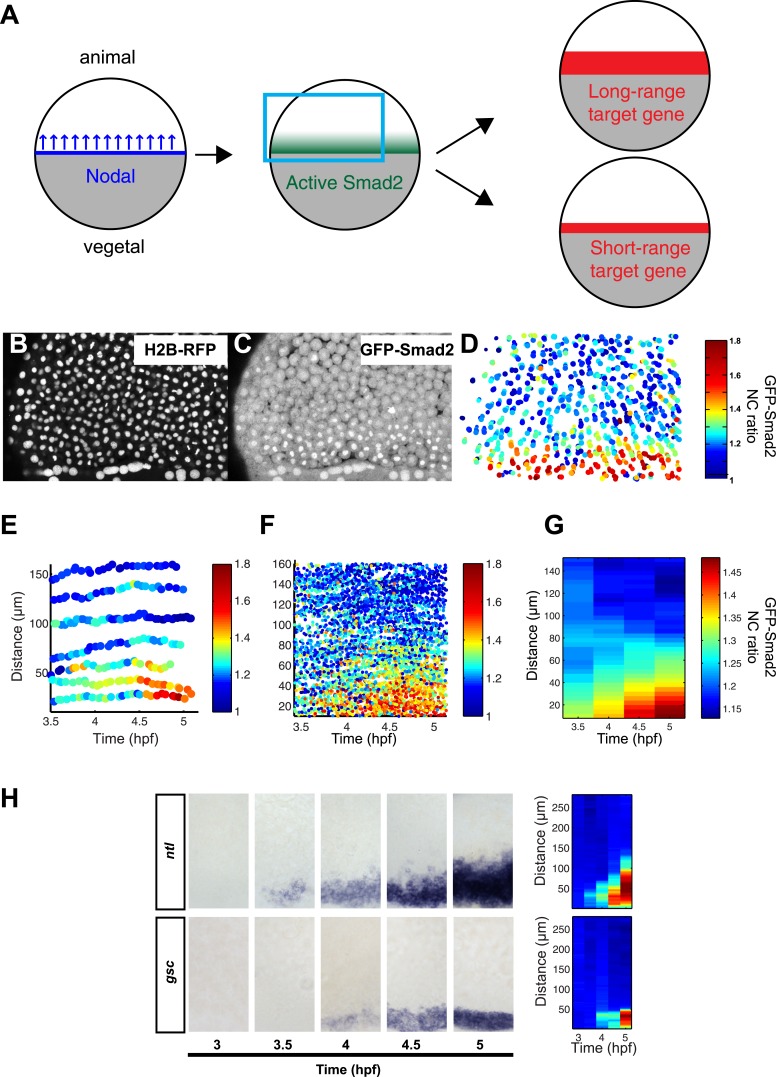
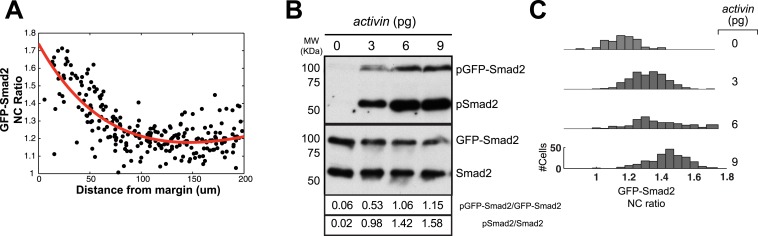
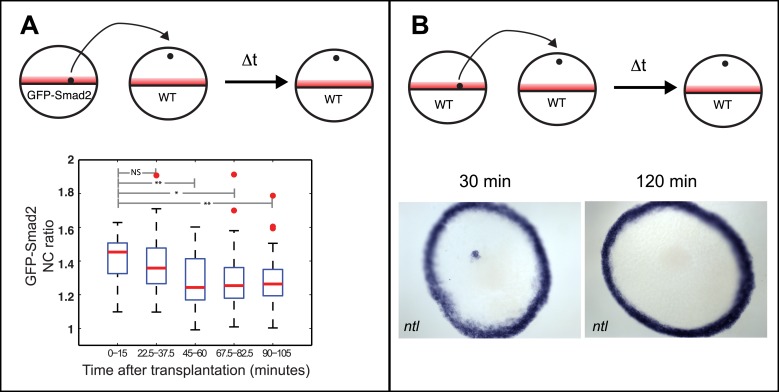
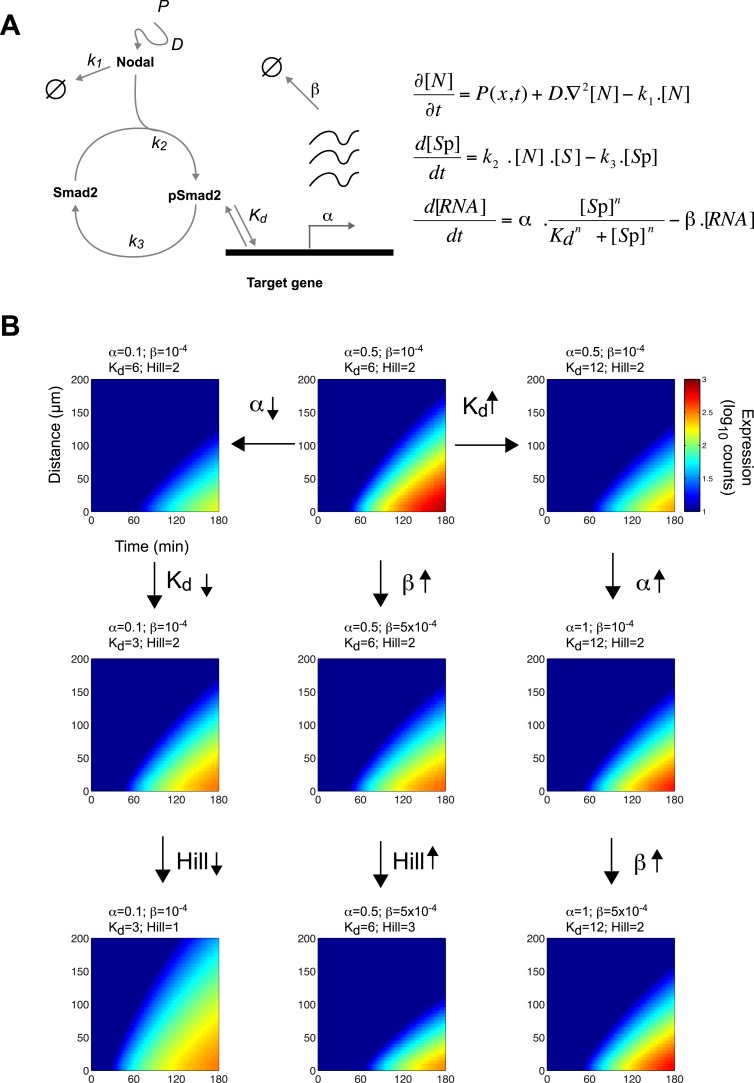
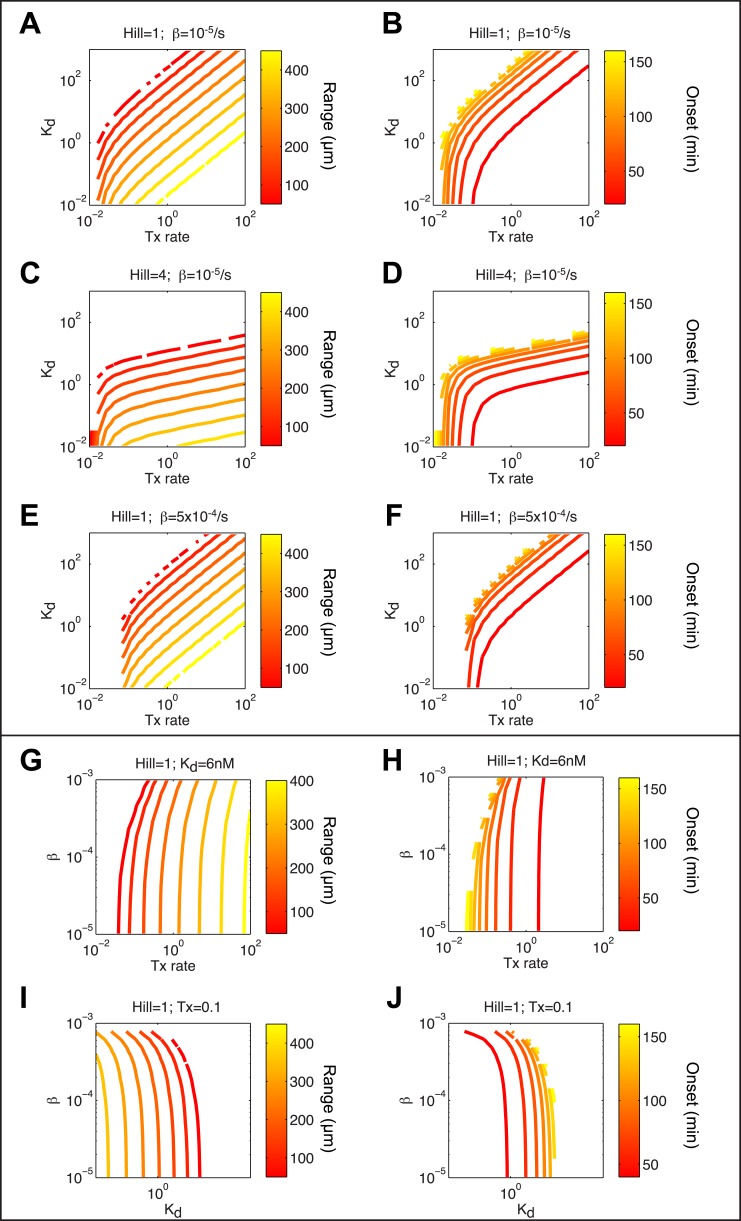
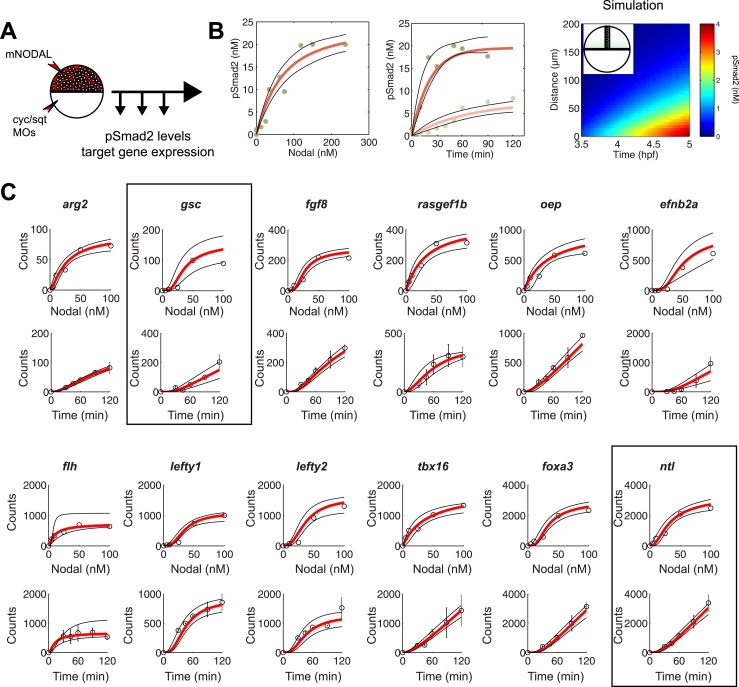
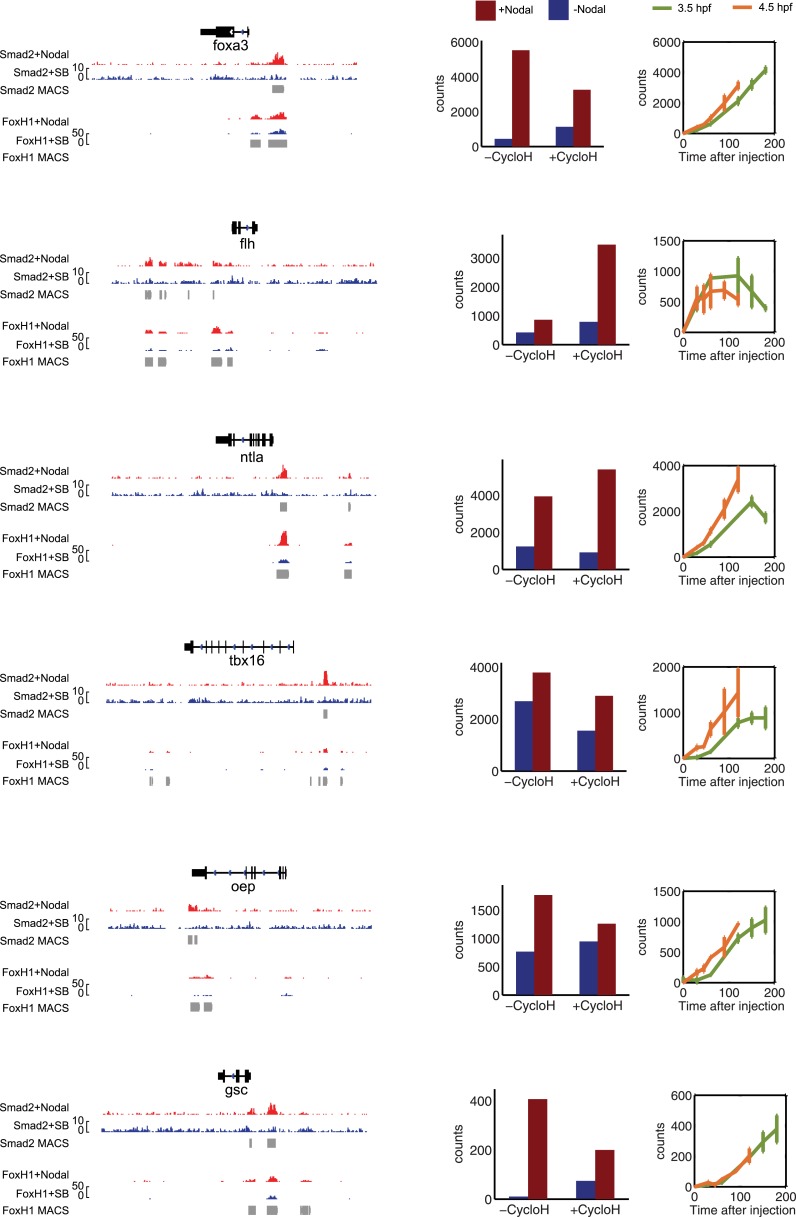
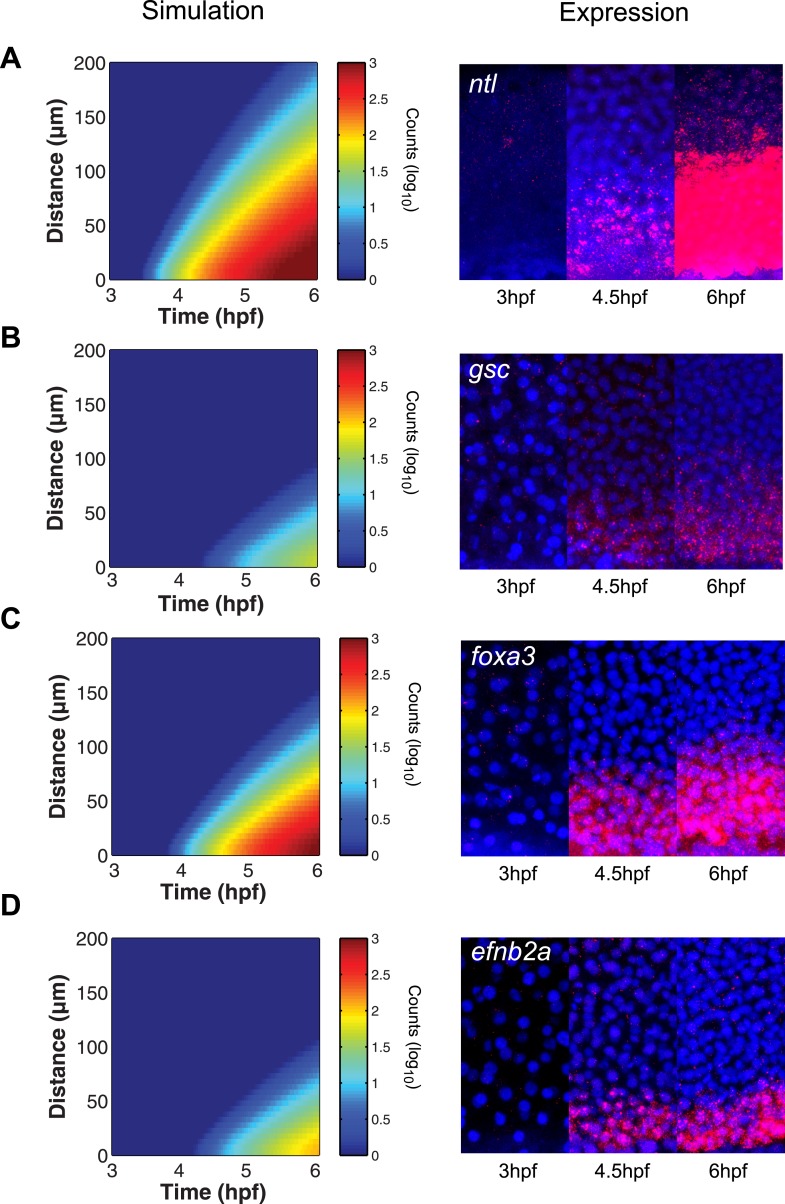
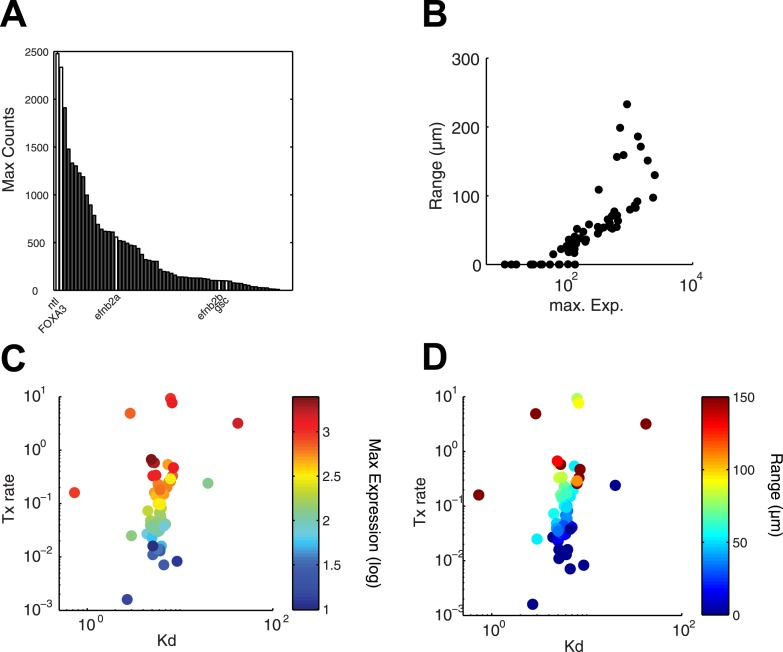
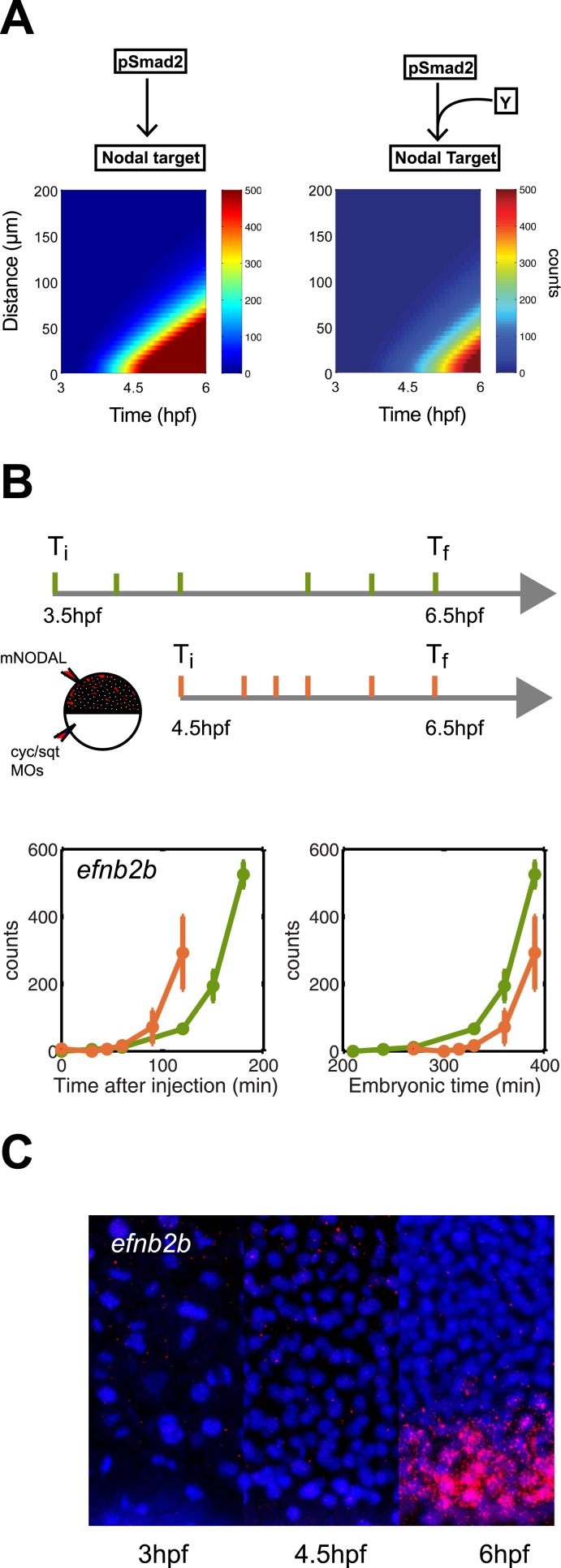
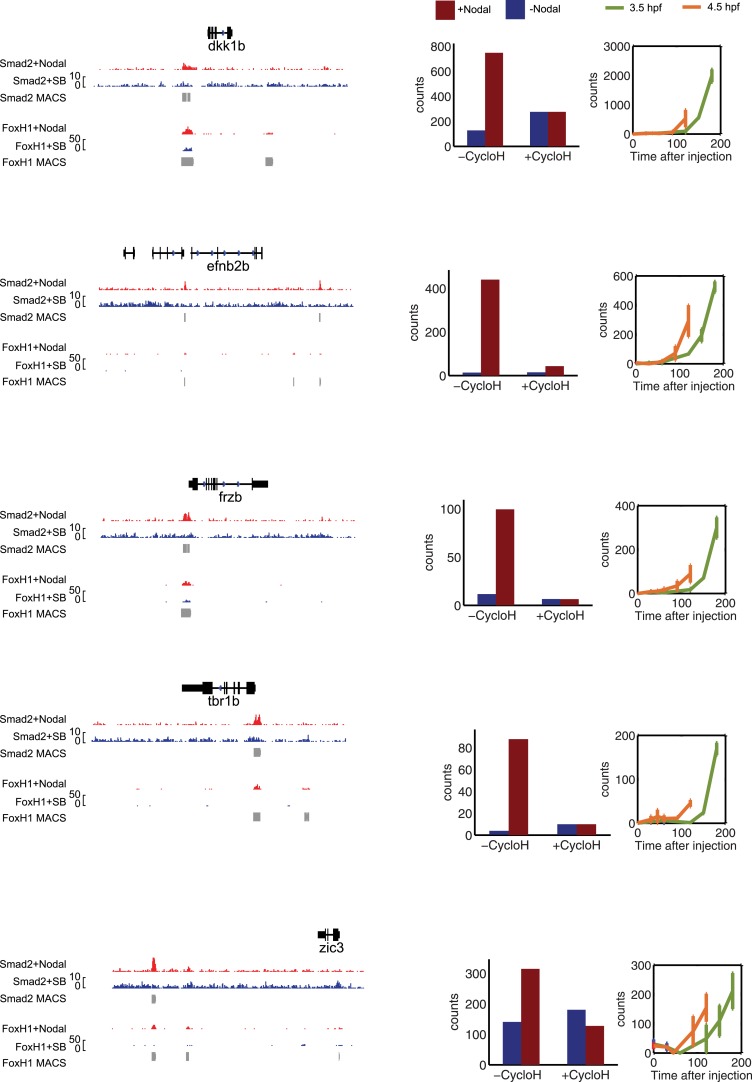
Morphogen gradients expose cells to different signal concentrations and induce target genes with different ranges of expression. To determine how the Nodal morphogen gradient induces distinct gene expression patterns during zebrafish embryogenesis, we measured the activation dynamics of the signal transducer Smad2 and the expression kinetics of long- and short-range target genes. We found that threshold models based on ligand concentration are insufficient to predict the response of target genes. Instead, morphogen interpretation is shaped by the kinetics of target gene induction: the higher the rate of transcription and the earlier the onset of induction, the greater the spatial range of expression. Thus, the timing and magnitude of target gene expression can be used to modulate the range of expression and diversify the response to morphogen gradients.
形态发生素梯度使细胞暴露于不同的信号浓度,并诱导具有不同表达范围的靶基因。为了确定Nodal形态发生素梯度如何在斑马鱼胚胎发育过程中诱导不同的基因表达模式,我们测量了信号转导子Smad2的激活动力学以及长程和短程靶基因的表达动力学。我们发现基于配体浓度的阈值模型不足以预测靶基因的反应。相反,形态发生素的解读由靶基因诱导的动力学决定:转录速率越高,诱导开始越早,表达的空间范围就越大。因此,靶基因表达的时间和幅度可用于调节表达范围,并使对形态发生素梯度的反应多样化。