Zhao Yunjie, Zeng Chen, Massiah Michael A
Department of Physics, The George Washington University, Washington, District of Columbia, United States of America.
Department of Physics, The George Washington University, Washington, District of Columbia, United States of America; Department of Physics, Huazhong University of Science and Technology, Wuhan, Hubei, China.
PLoS One. 2015 Apr 13;10(4):e0124377. doi: 10.1371/journal.pone.0124377. eCollection 2015.
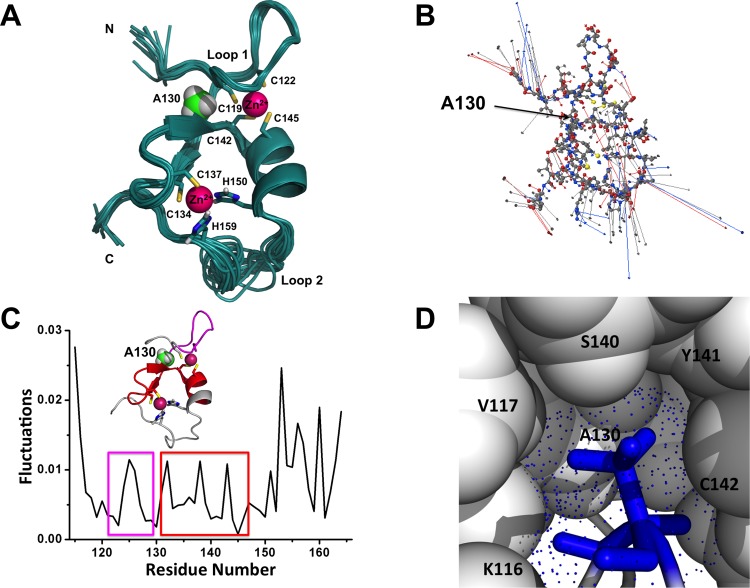
The zinc-binding Bbox1 domain in protein MID1, a member of the TRIM family of proteins, facilitates the ubiquitination of the catalytic subunit of protein phosphatase 2A and alpha4, a protein regulator of PP2A. The natural mutation of residue A130 to a valine or threonine disrupts substrate recognition and catalysis. While NMR data revealed the A130T mutant Bbox1 domain failed to coordinate both structurally essential zinc ions and resulted in an unfolded structure, the unfolding mechanism is unknown. Principle component analysis revealed that residue A130 served as a hinge point between the structured β-strand-turn-β-strand (β-turn-β) and the lasso-like loop sub-structures that constitute loop1 of the ββα-RING fold that the Bbox1 domain adopts. Backbone RMSD data indicate significant flexibility and departure from the native structure within the first 5 ns of the molecular dynamics (MD) simulation for the A130V mutant (>6 Å) and after 30 ns for A130T mutant (>6 Å). Overall RMSF values were higher for the mutant structures and showed increased flexibility around residues 125 and 155, regions with zinc-coordinating residues. Simulated pKa values of the sulfhydryl group of C142 located near A130 suggested an increased in value to ~9.0, paralleling the increase in the apparent dielectric constants for the small cavity near residue A130. Protonation of the sulfhydryl group would disrupt zinc-coordination, directly contributing to unfolding of the Bbox1. Together, the increased motion of residues of loop 1, which contains four of the six zinc-binding cysteine residues, and the increased pKa of C142 could destabilize the structure of the zinc-coordinating residues and contribute to the unfolding.
蛋白质MID1是TRIM蛋白家族的成员之一,其锌结合Bbox1结构域促进蛋白磷酸酶2A催化亚基和α4(一种PP2A的蛋白质调节剂)的泛素化。残基A130自然突变为缬氨酸或苏氨酸会破坏底物识别和催化作用。虽然核磁共振数据显示A130T突变体Bbox1结构域无法配位两个结构必需的锌离子并导致结构展开,但其展开机制尚不清楚。主成分分析表明,残基A130是构成Bbox1结构域所采用的ββα-RING折叠环1的结构化β-链-转角-β-链(β-转角-β)和套索样环子结构之间的铰链点。主链均方根偏差数据表明,A130V突变体在分子动力学(MD)模拟的前5 ns内(>6 Å)以及A130T突变体在30 ns后(>6 Å)具有显著的灵活性并偏离天然结构。突变体结构的整体均方根波动值更高,并且在残基125和155周围显示出增加的灵活性,这两个区域是锌配位残基所在区域。位于A130附近的C142巯基的模拟pKa值表明其值增加到约9.0,这与残基A130附近小腔的表观介电常数增加平行。巯基的质子化会破坏锌配位,直接导致Bbox1展开。总之,包含六个锌结合半胱氨酸残基中的四个的环1残基的运动增加以及C142 pKa的增加可能会使锌配位残基的结构不稳定并导致展开。