Agarwal Rachna, Hasan S Saif, Jones LaDonna M, Stofleth Jason T, Ryan Christopher M, Whitelegge Julian P, Kehoe David M, Cramer William A
†Department of Biological Sciences, Purdue University, West Lafayette, Indiana 47907, United States.
‡Department of Biology, Indiana University, Bloomington, Indiana 47405, United States.
Biochemistry. 2015 May 26;54(20):3151-63. doi: 10.1021/acs.biochem.5b00279. Epub 2015 May 12.
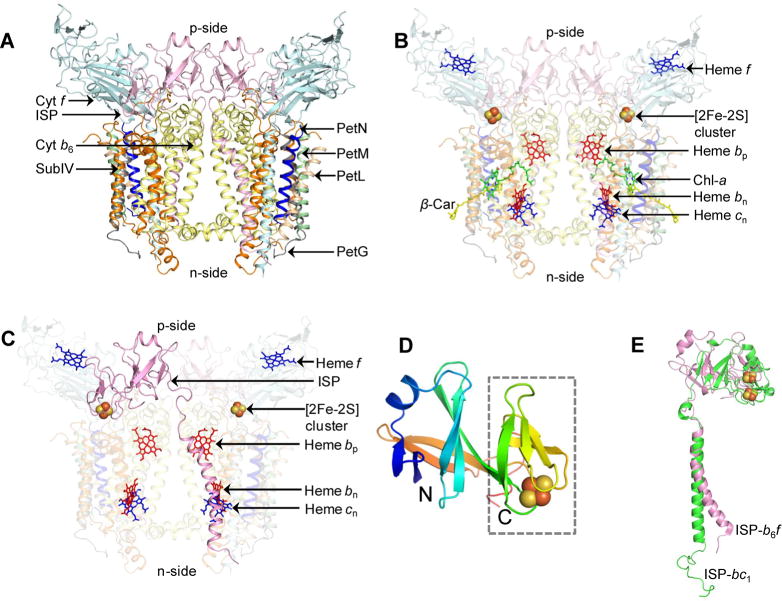
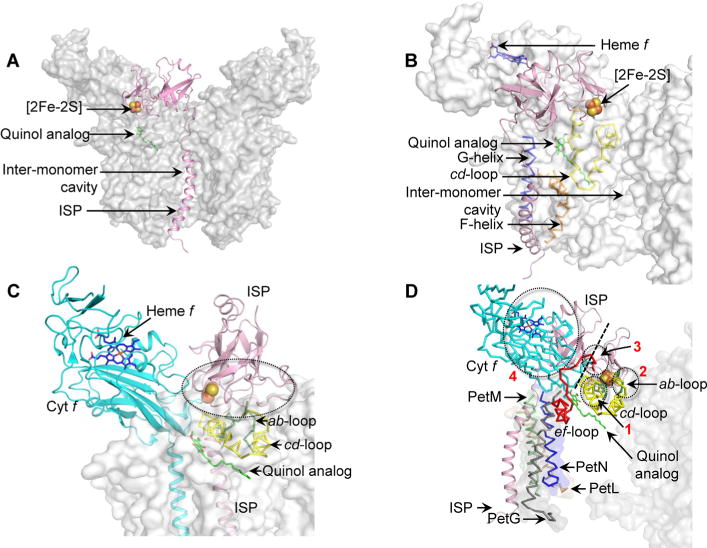
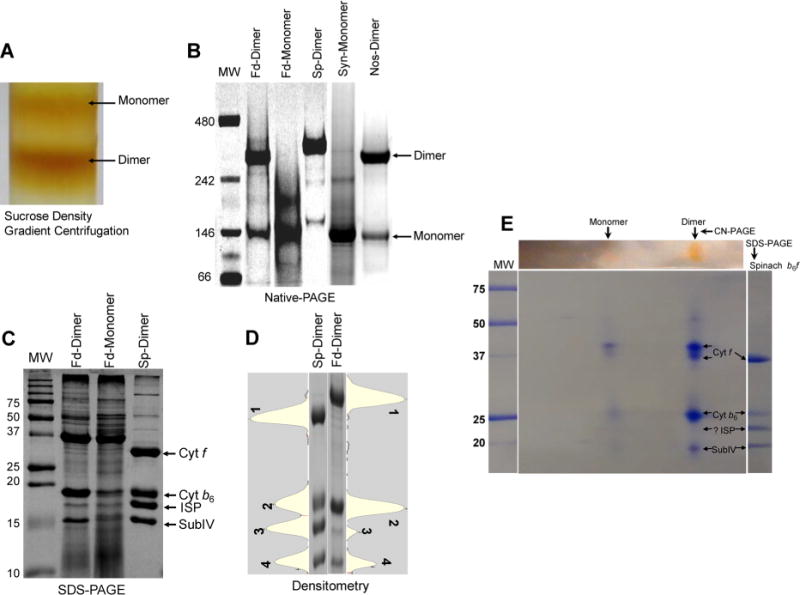
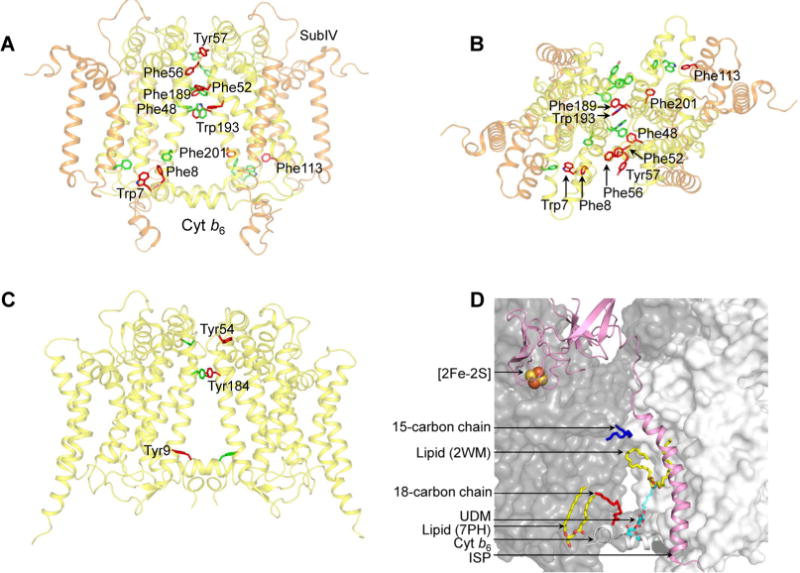
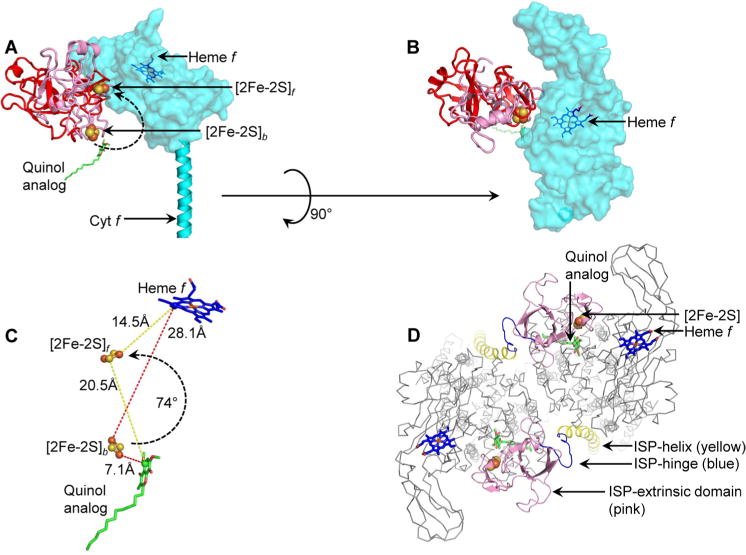
Domain swapping that contributes to the stability of biologically crucial multisubunit complexes has been implicated in protein oligomerization. In the case of membrane protein assemblies, domain swapping of the iron-sulfur protein (ISP) subunit occurs in the hetero-oligomeric cytochrome b6f and bc1 complexes, which are organized as symmetric dimers that generate the transmembrane proton electrochemical gradient utilized for ATP synthesis. In these complexes, the ISP C-terminal predominantly β-sheet extrinsic domain containing the redox-active [2Fe-2S] cluster resides on the electrochemically positive side of each monomer in the dimeric complex. This domain is bound to the membrane sector of the complex through an N-terminal transmembrane α-helix that is "swapped' to the other monomer of the complex where it spans the complex and the membrane. Detailed analysis of the function and structure of the b6f complex isolated from the cyanobacterium Fremyella diplosiphon SF33 shows that the domain-swapped ISP structure is necessary for function but is not necessarily essential for maintenance of the dimeric structure of the complex. On the basis of crystal structures of the cytochrome complex, the stability of the cytochrome dimer is attributed to specific intermonomer protein-protein and protein-lipid hydrophobic interactions. The geometry of the domain-swapped ISP structure is proposed to be a consequence of the requirement that the anchoring helix of the ISP not perturb the heme organization or quinone channel in the conserved core of each monomer.
有助于生物关键多亚基复合物稳定性的结构域交换与蛋白质寡聚化有关。就膜蛋白组装而言,铁硫蛋白(ISP)亚基的结构域交换发生在异源寡聚细胞色素b6f和bc1复合物中,这些复合物以对称二聚体形式组织,产生用于ATP合成的跨膜质子电化学梯度。在这些复合物中,含有氧化还原活性[2Fe-2S]簇的ISP C末端主要为β折叠的外在结构域位于二聚体复合物中每个单体的电化学正侧。该结构域通过一个N末端跨膜α螺旋与复合物的膜部分结合,该α螺旋“交换”到复合物的另一个单体上,跨越复合物和膜。对从蓝藻双孢 Fremyella diplosiphon SF33 分离的 b6f 复合物的功能和结构进行的详细分析表明,结构域交换的 ISP 结构对功能是必要的,但不一定对维持复合物的二聚体结构至关重要。根据细胞色素复合物的晶体结构,细胞色素二聚体的稳定性归因于特定的单体间蛋白质-蛋白质和蛋白质-脂质疏水相互作用。有人提出,结构域交换的 ISP 结构的几何形状是由于 ISP 的锚定螺旋不能干扰每个单体保守核心中的血红素组织或醌通道这一要求所致。