Vafabakhsh Reza, Levitz Joshua, Isacoff Ehud Y
Department of Molecular and Cell Biology, University of California, Berkeley, California 94720, USA.
Helen Wills Neuroscience Institute, University of California, Berkeley, California 94720, USA.
Nature. 2015 Aug 27;524(7566):497-501. doi: 10.1038/nature14679. Epub 2015 Aug 10.
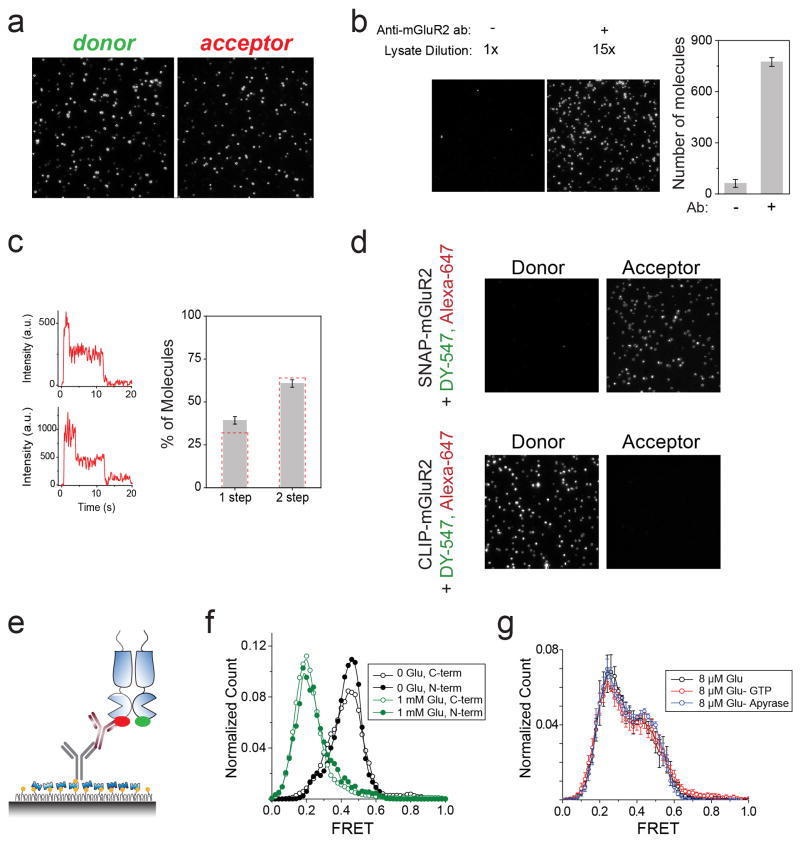
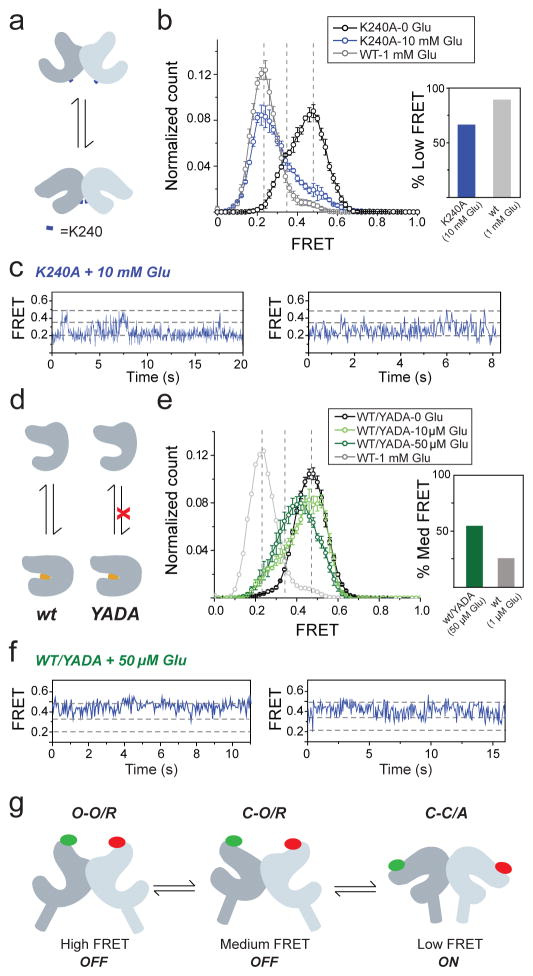
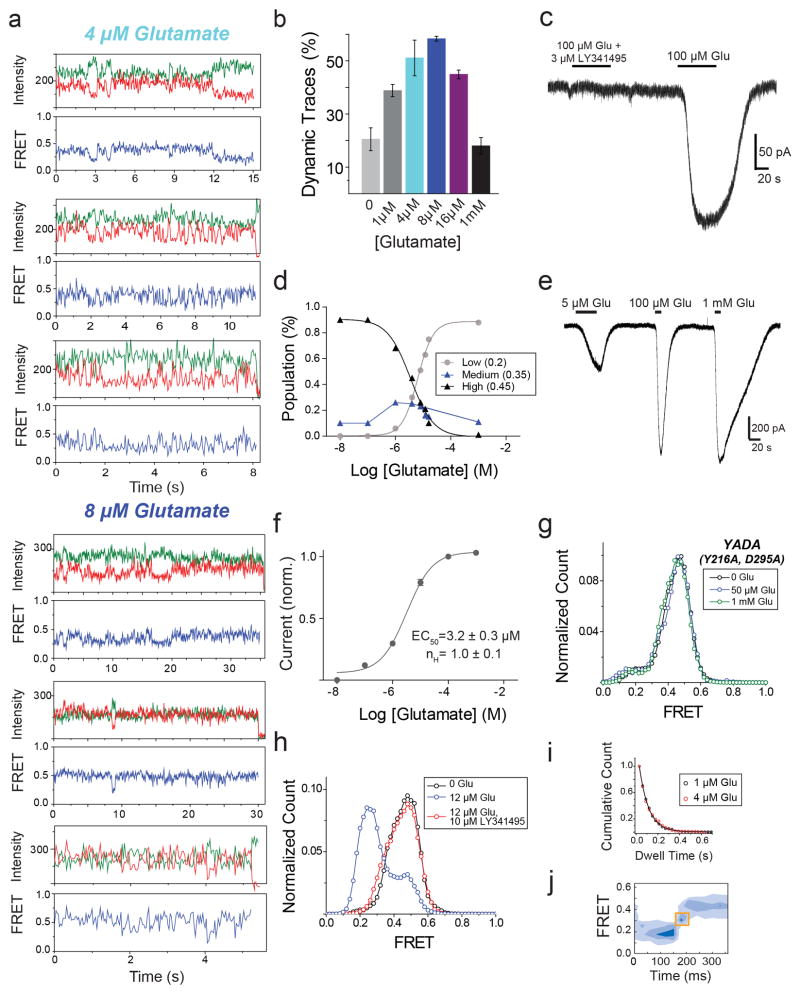
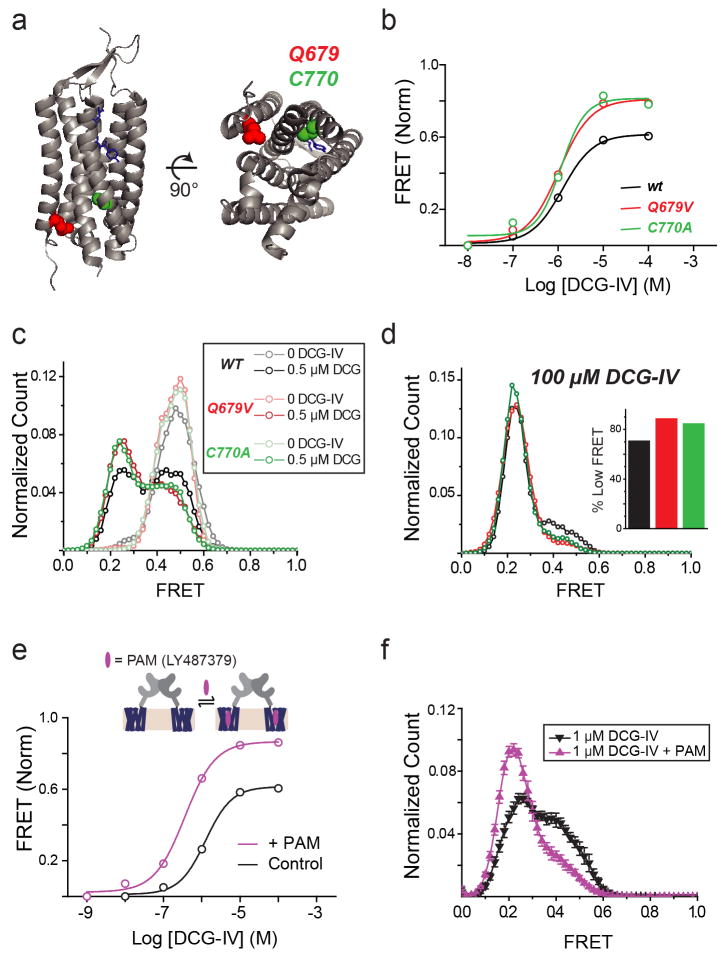
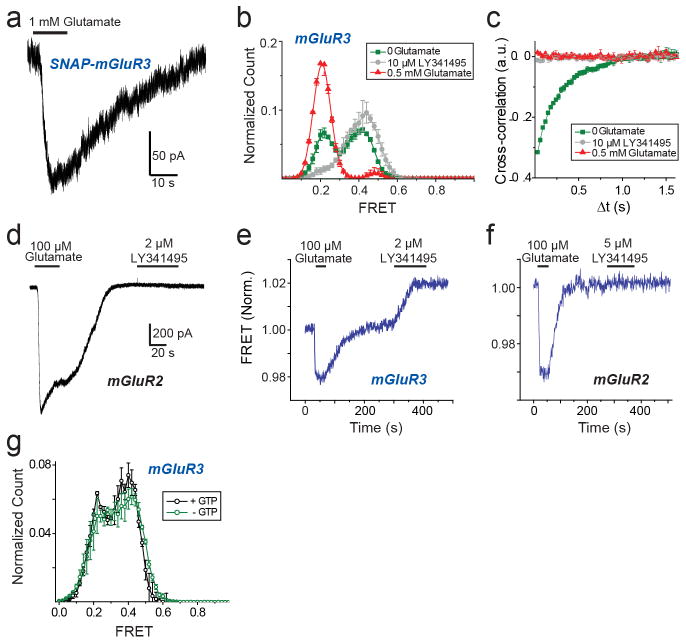
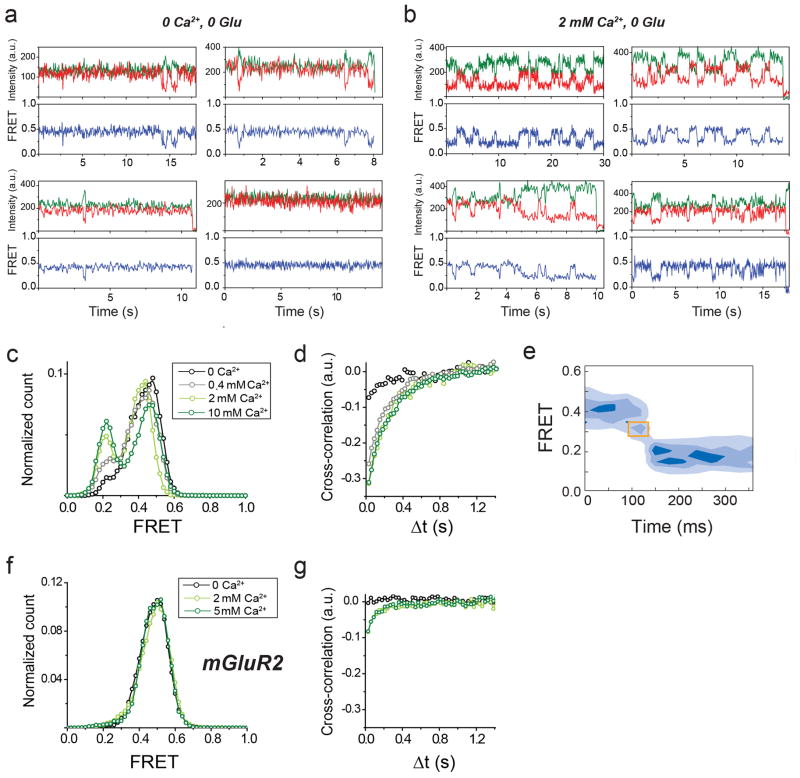
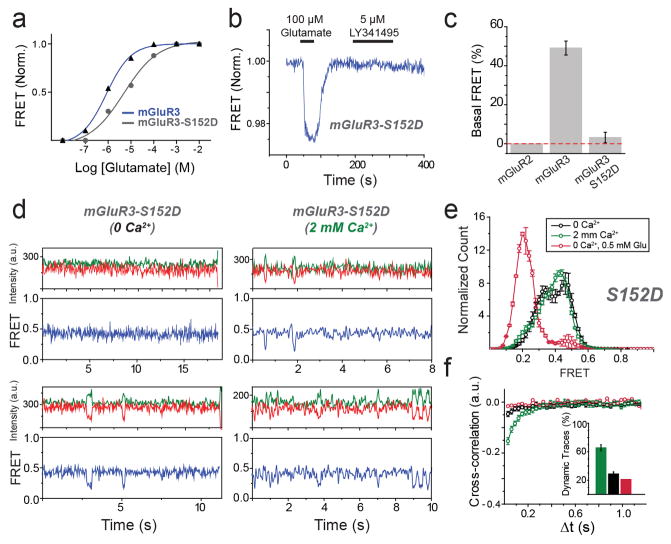
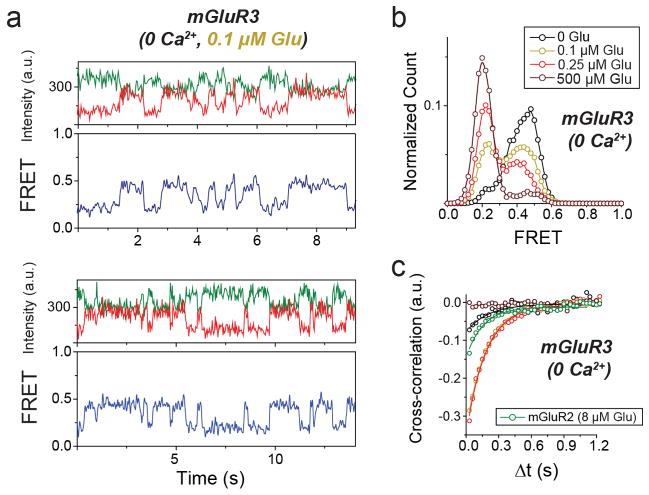
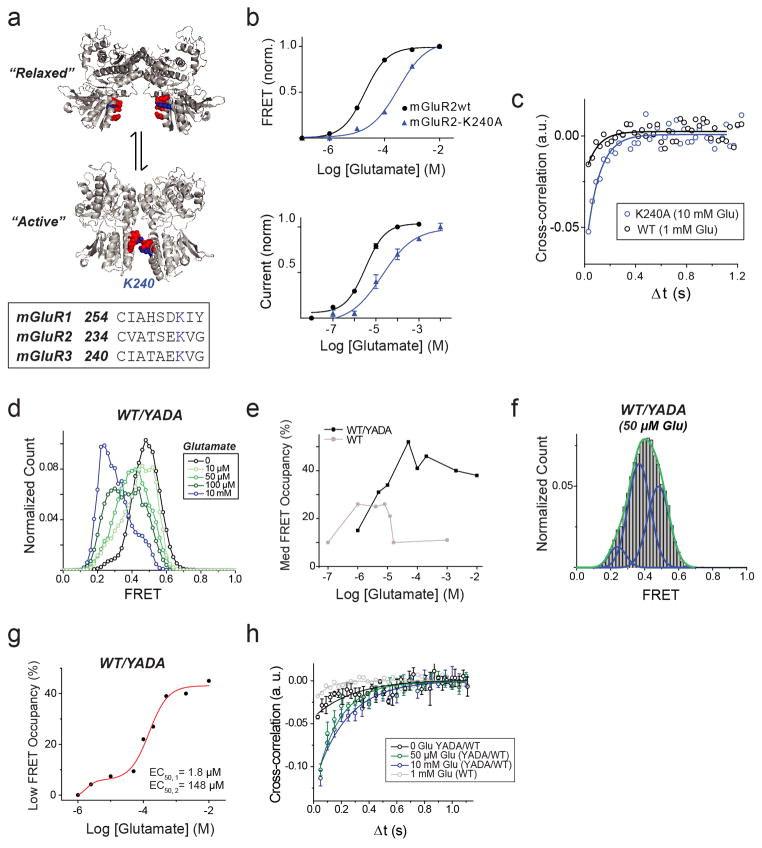
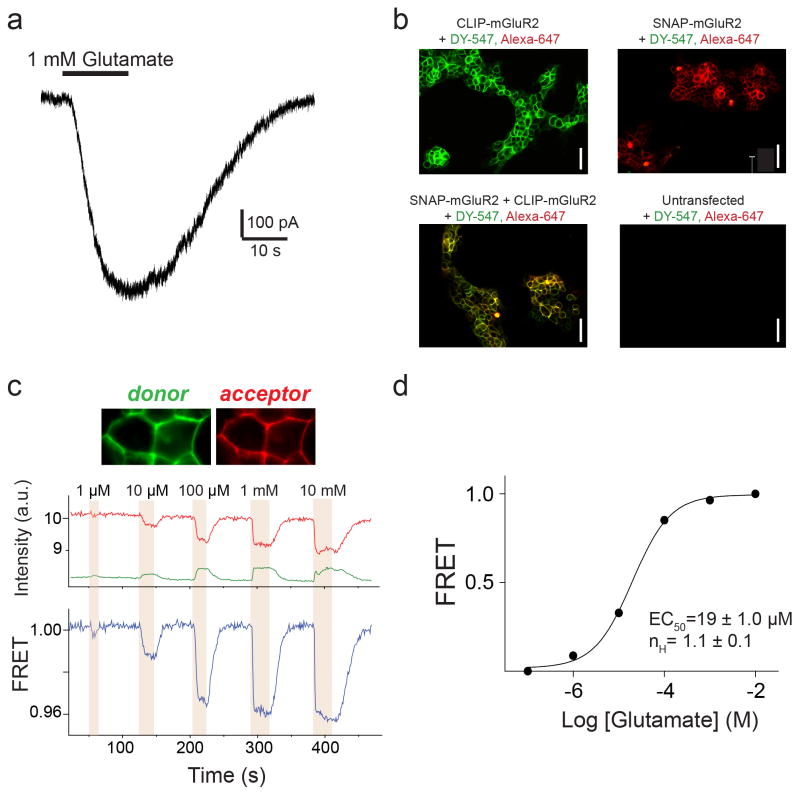
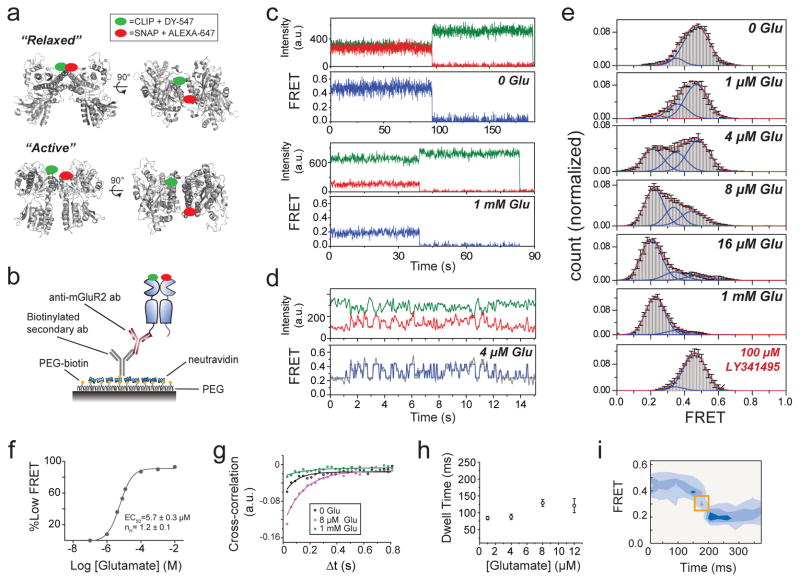
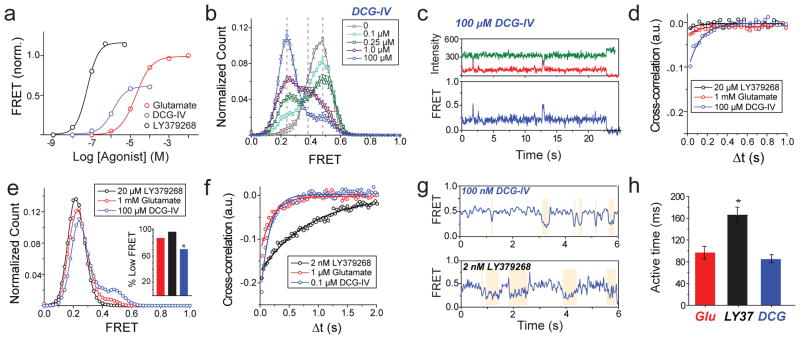
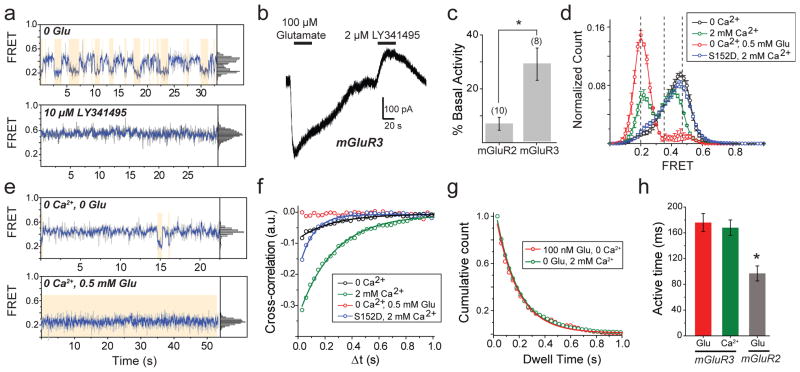
G-protein-coupled receptors (GPCRs) constitute the largest family of membrane receptors in eukaryotes. Crystal structures have provided insight into GPCR interactions with ligands and G proteins, but our understanding of the conformational dynamics of activation is incomplete. Metabotropic glutamate receptors (mGluRs) are dimeric class C GPCRs that modulate neuronal excitability, synaptic plasticity, and serve as drug targets for neurological disorders. A 'clamshell' ligand-binding domain (LBD), which contains the ligand-binding site, is coupled to the transmembrane domain via a cysteine-rich domain, and LBD closure seems to be the first step in activation. Crystal structures of isolated mGluR LBD dimers led to the suggestion that activation also involves a reorientation of the dimer interface from a 'relaxed' to an 'active' state, but the relationship between ligand binding, LBD closure and dimer interface rearrangement in activation remains unclear. Here we use single-molecule fluorescence resonance energy transfer to probe the activation mechanism of full-length mammalian group II mGluRs. We show that the LBDs interconvert between three conformations: resting, activated and a short-lived intermediate state. Orthosteric agonists induce transitions between these conformational states, with efficacy determined by occupancy of the active conformation. Unlike mGluR2, mGluR3 displays basal dynamics, which are Ca(2+)-dependent and lead to basal protein activation. Our results support a general mechanism for the activation of mGluRs in which agonist binding induces closure of the LBDs, followed by dimer interface reorientation. Our experimental strategy should be widely applicable to study conformational dynamics in GPCRs and other membrane proteins.
G蛋白偶联受体(GPCRs)是真核生物中最大的膜受体家族。晶体结构为深入了解GPCR与配体及G蛋白的相互作用提供了线索,但我们对激活过程中的构象动力学的理解仍不完整。代谢型谷氨酸受体(mGluRs)是二聚体C类GPCR,可调节神经元兴奋性、突触可塑性,并作为神经疾病的药物靶点。一个包含配体结合位点的“蛤壳”样配体结合结构域(LBD)通过富含半胱氨酸的结构域与跨膜结构域相连,LBD的闭合似乎是激活的第一步。分离的mGluR LBD二聚体的晶体结构表明,激活还涉及二聚体界面从“松弛”状态到“活性”状态的重新定向,但配体结合、LBD闭合与激活过程中二聚体界面重排之间的关系仍不清楚。在此,我们使用单分子荧光共振能量转移来探究全长哺乳动物II组mGluRs的激活机制。我们发现LBDs在三种构象之间相互转换:静息态、激活态和一种短暂的中间态。正构激动剂诱导这些构象状态之间的转变,其效力由活性构象的占有率决定。与mGluR2不同,mGluR3表现出基础动力学,这是Ca(2+)依赖性的,并导致基础蛋白激活。我们的结果支持一种mGluRs激活的通用机制,即激动剂结合诱导LBDs闭合,随后二聚体界面重新定向。我们的实验策略应广泛适用于研究GPCRs和其他膜蛋白中的构象动力学。