Department of Geology and PalaeontologyNational Museum of Nature and ScienceTsukuba, Japan; Department of GeologyUniversity of OtagoDunedin, New Zealand.
Department of Geology University of Otago Dunedin, New Zealand.
R Soc Open Sci. 2015 Dec 2;2(12):150476. doi: 10.1098/rsos.150476. eCollection 2015 Dec.
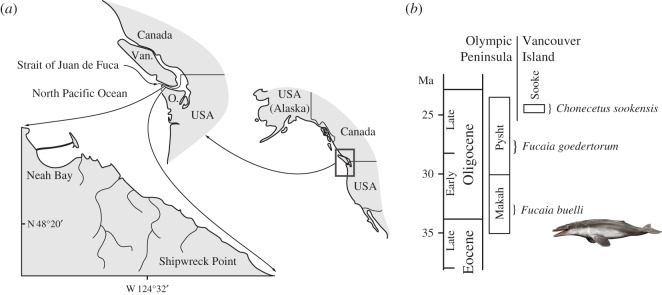
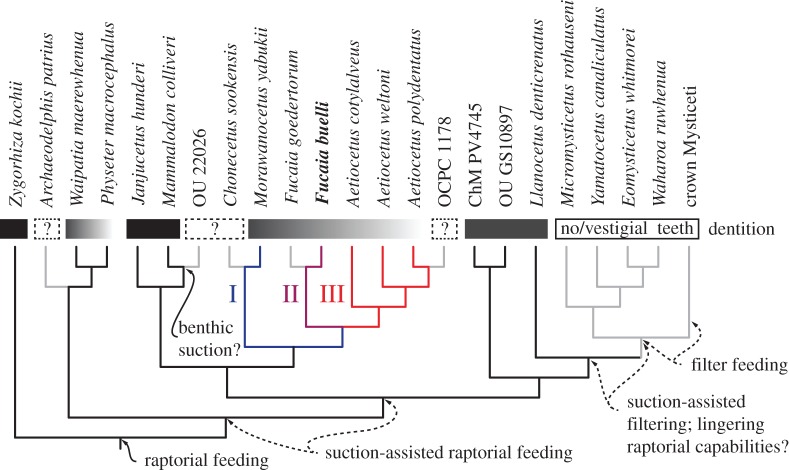
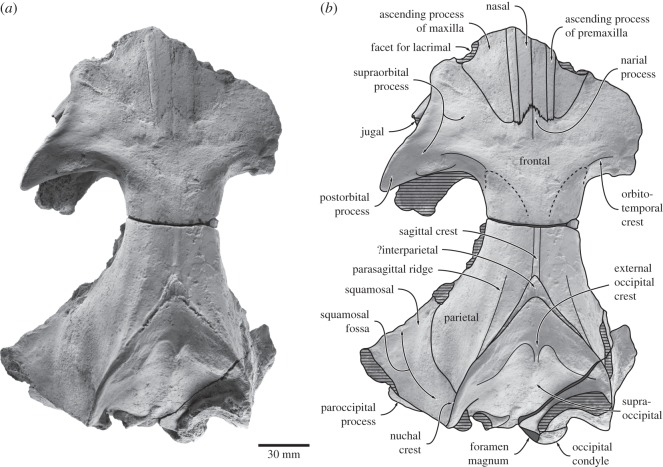
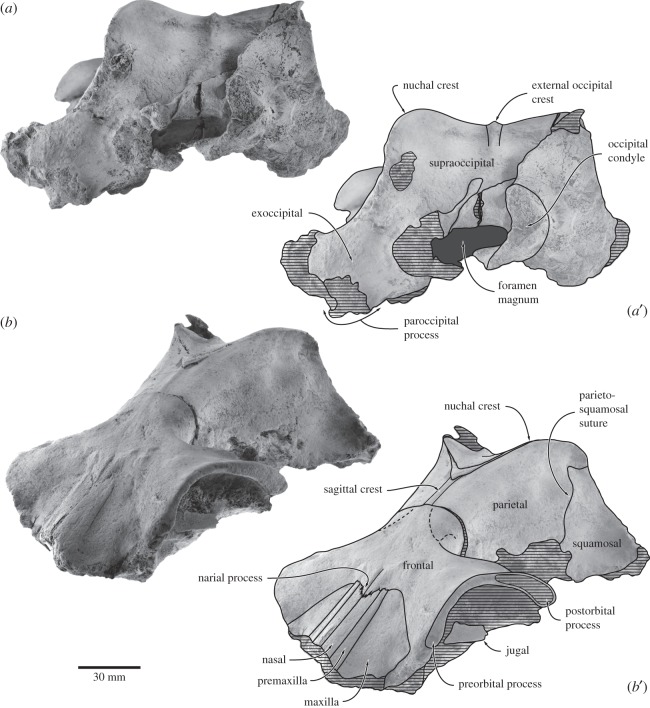
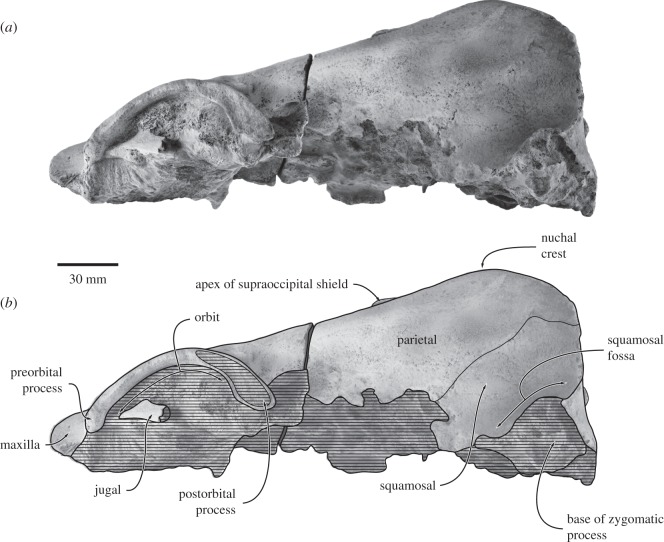
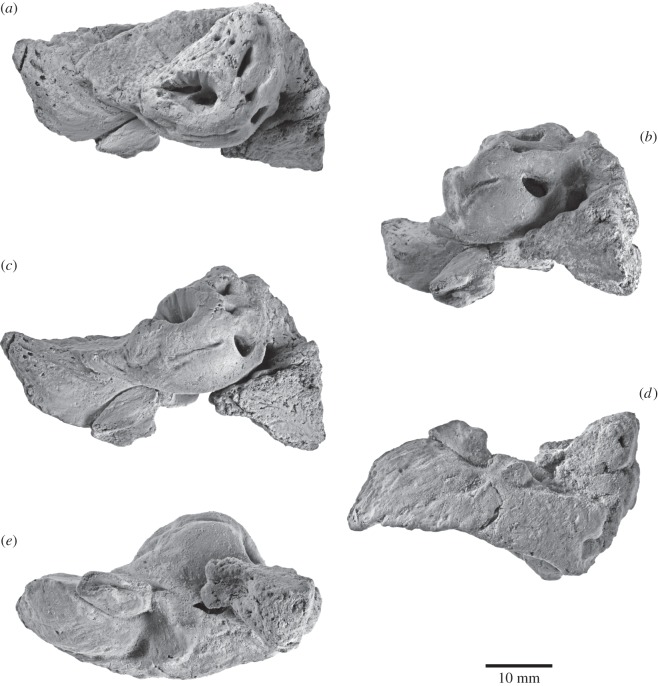
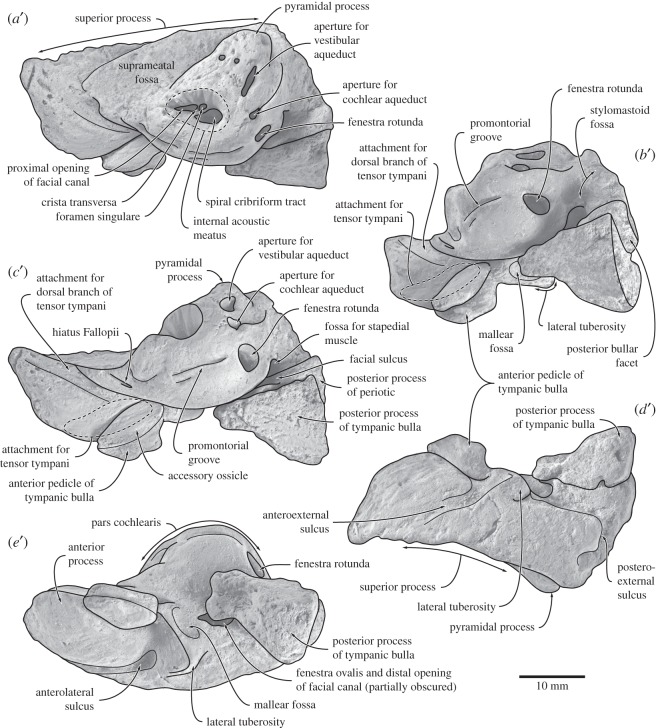
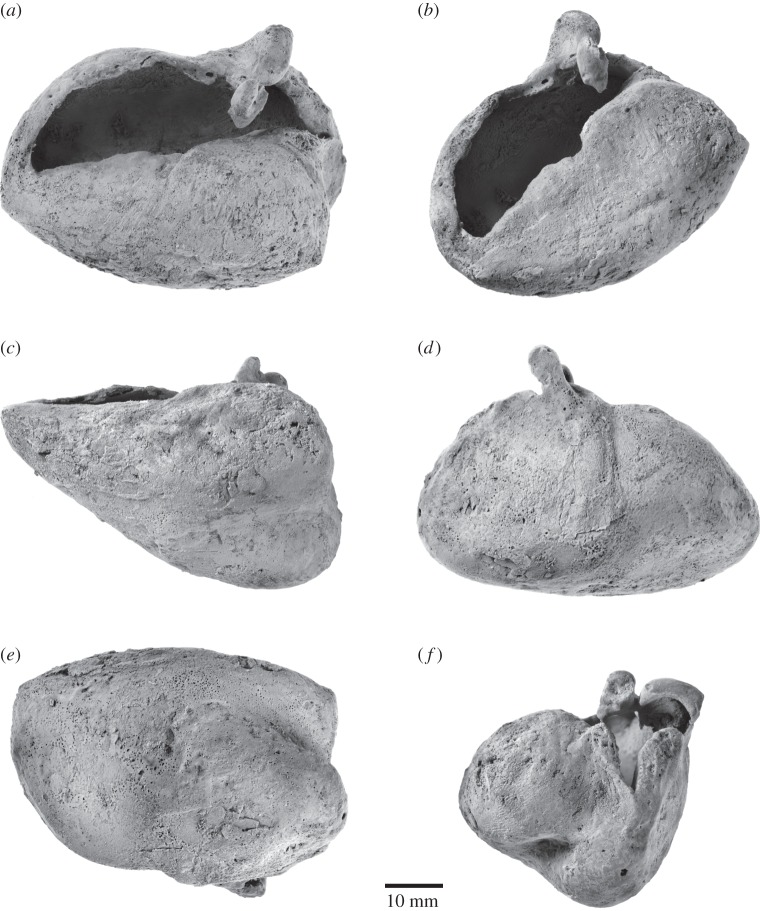
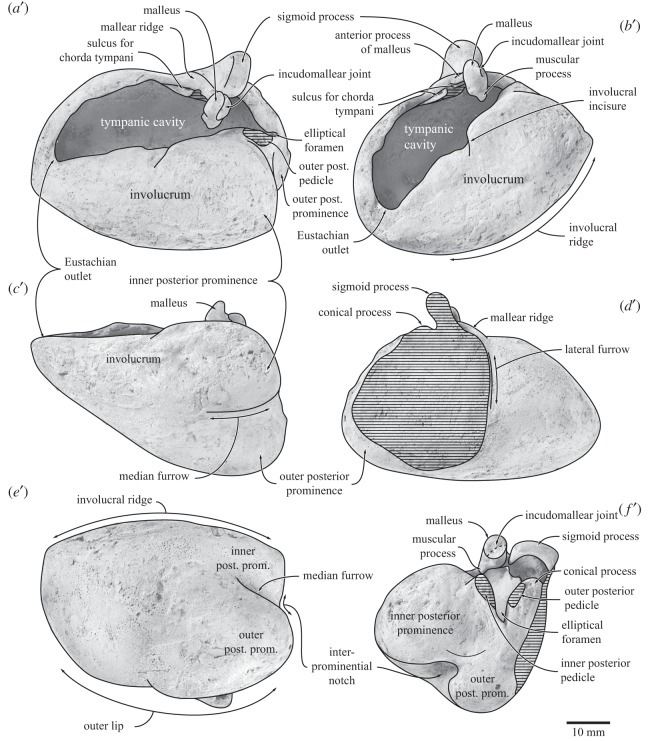
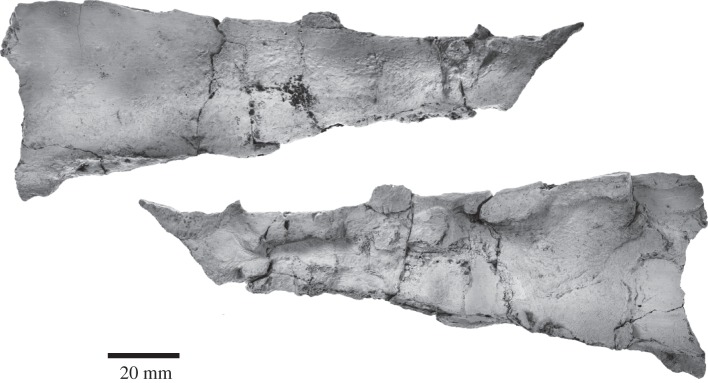
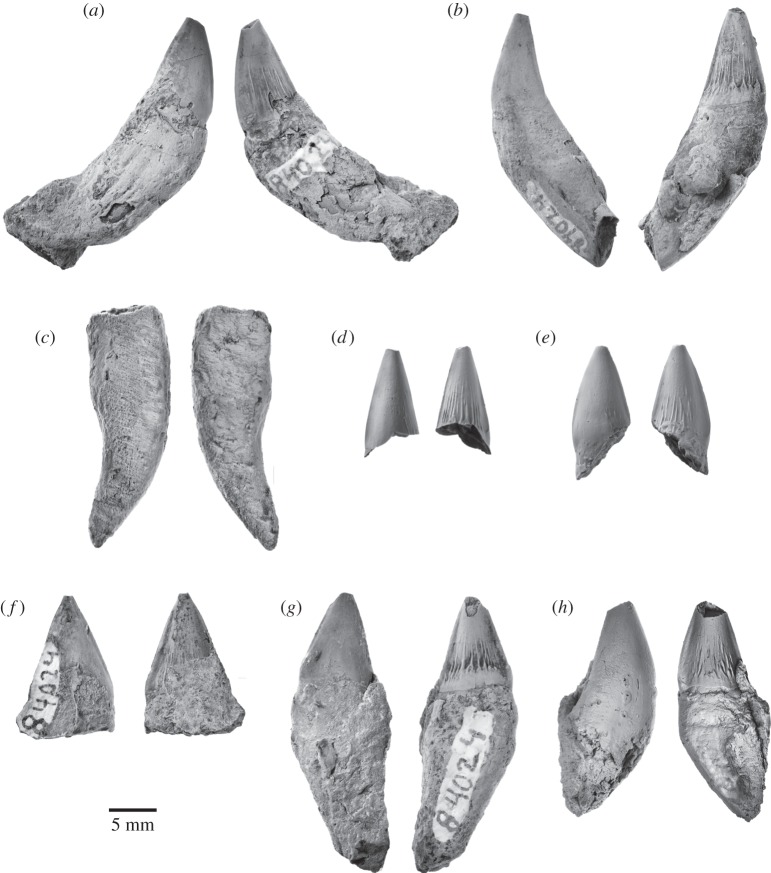
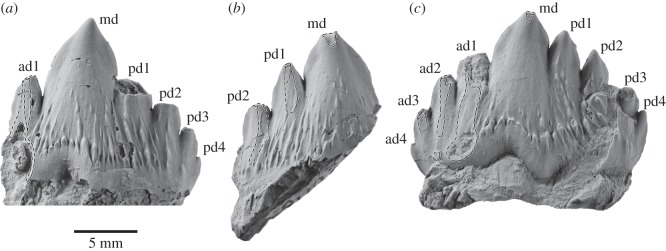
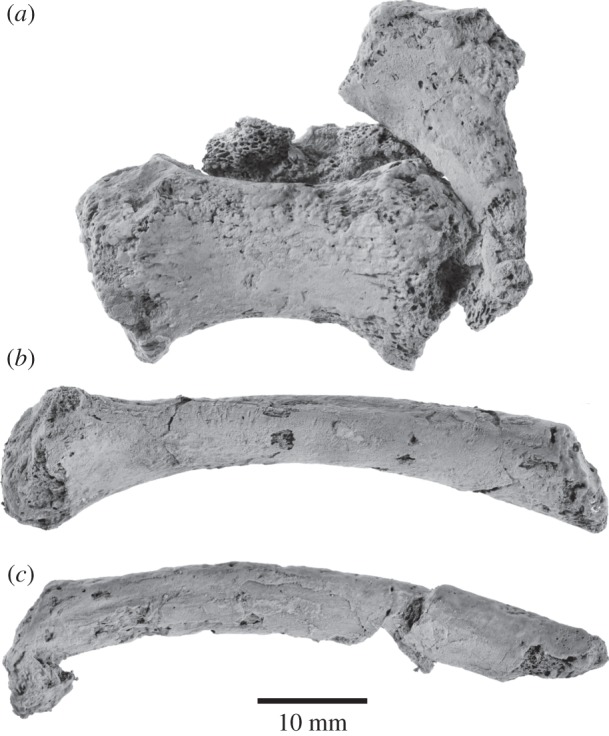
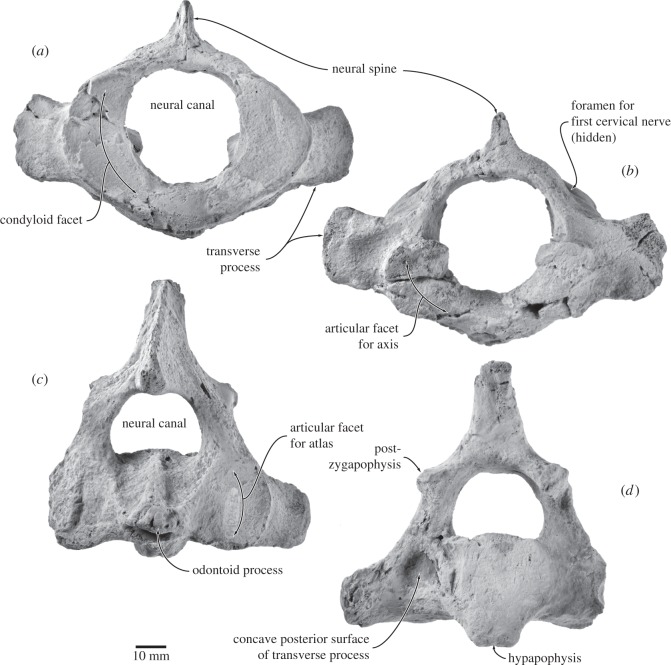
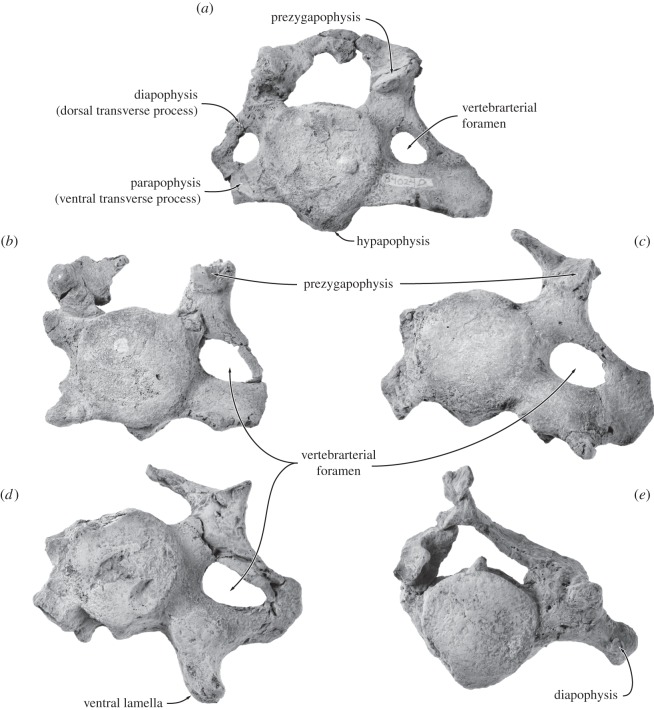
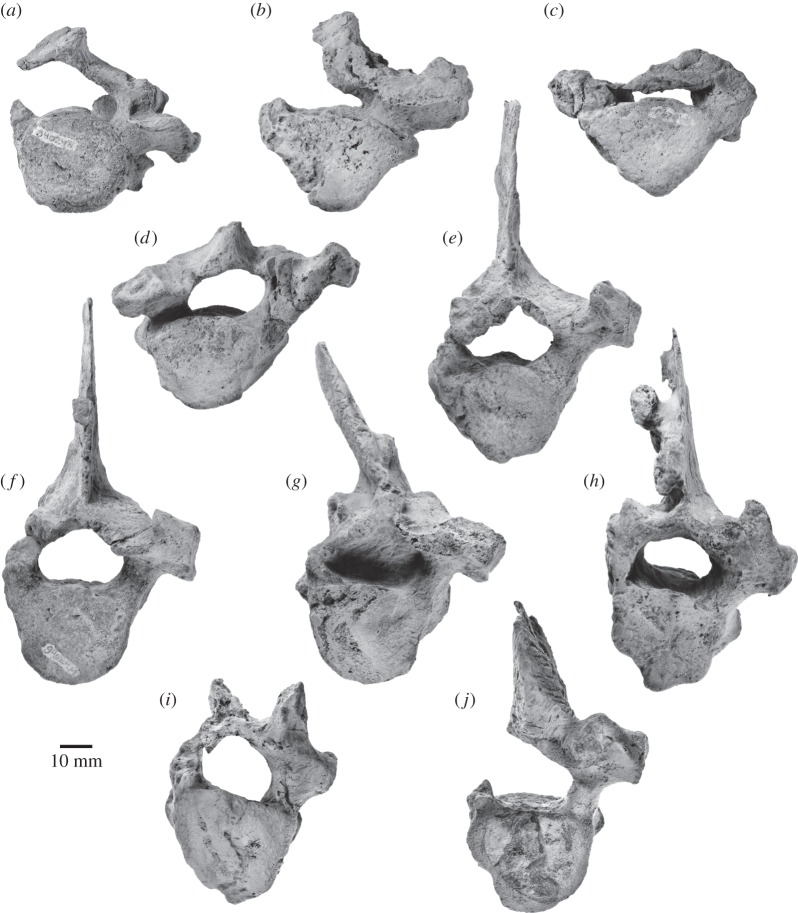
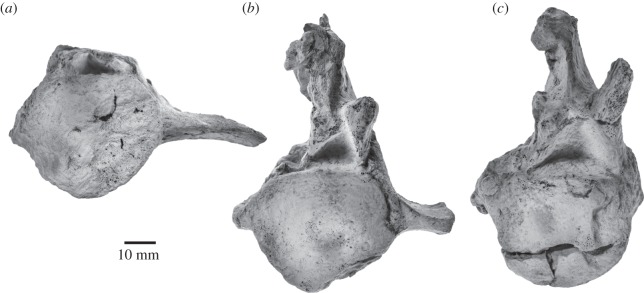
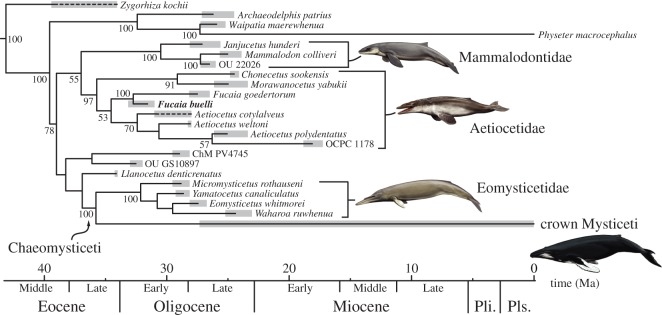
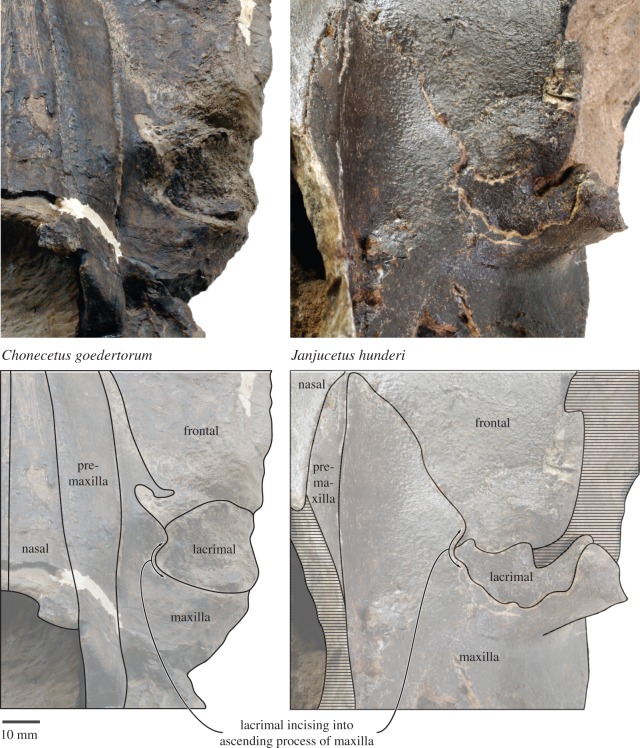
Archaic toothed mysticetes represent the evolutionary transition from raptorial to bulk filter feeding in baleen whales. Aetiocetids, in particular, preserve an intermediate morphological stage in which teeth functioned alongside a precursor of baleen, the hallmark of all modern mysticetes. To date, however, aetiocetids are almost exclusively Late Oligocene and coeval with both other toothed mysticetes and fully fledged filter feeders. By contrast, reports of cetaceans from the Early Oligocene remain rare, leaving the origins of aetiocetids, and thus of baleen, largely in the dark. Here, we report a new aetiocetid, Fucaia buelli, from the earliest Oligocene (ca 33-31 Ma) of western North America. The new material narrows the temporal gap between aetiocetids and the oldest known mysticete, Llanocetus (ca 34 Ma). The specimen preserves abundant morphological detail relating to the phylogenetically informative ear bones (otherwise poorly documented in this family), the hyoid apparatus and much of the (heterodont) dentition. Fucaia comprises some of the smallest known mysticetes, comparable in size with the smallest odontocetes. Based on their phylogenetic relationships and dental and mandibular morphology, including tooth wear patterns, we propose that aetiocetids were suction-assisted raptorial feeders and interpret this strategy as a crucial, intermediary step, enabling the transition from raptorial to filter feeding. Following this line of argument, a combination of raptorial and suction feeding would have been ancestral to all toothed mysticetes, and possibly even baleen whales as a whole.
古生齿鲸代表了从捕食性到须鲸过滤进食的进化过渡。尤其是矛齿鲸,它们保留了一个中间形态阶段,在这个阶段中,牙齿与须鲸的前身——所有现代须鲸的标志——共同发挥作用。然而,迄今为止,矛齿鲸几乎仅存在于晚渐新世,与其他有齿的须鲸和完全成熟的滤食者同时存在。相比之下,早渐新世的鲸目动物的报告仍然很少,这使得矛齿鲸的起源,以及须鲸的起源,在很大程度上仍不清楚。在这里,我们报告了一种来自北美西部最早的渐新世(约 33-31 百万年前)的新的矛齿鲸属种,福凯亚·布埃利。新材料缩小了矛齿鲸和已知最古老的须鲸——兰诺鲸(约 34 百万年前)之间的时间差距。该标本保留了丰富的形态学细节,涉及到与系统发育相关的耳骨(在这个家族中记录不佳)、舌骨器官和大部分(异齿)牙齿。福凯亚属是已知最小的须鲸之一,与最小的齿鲸相当。基于它们的系统发育关系以及牙齿和下颌骨的形态,包括牙齿磨损模式,我们提出矛齿鲸是一种吸力辅助的捕食性动物,并将这种策略解释为一种至关重要的中间步骤,使从捕食性到滤食的过渡成为可能。根据这一论点,捕食性和吸力性进食的结合可能是所有有齿须鲸的祖先,甚至可能是整个须鲸目动物的祖先。