Nilsson Anders K, Fahlberg Per, Johansson Oskar N, Hamberg Mats, Andersson Mats X, Ellerström Mats
Department of Biological and Environmental Sciences, University of Gothenburg, Box 461, SE-405 30 Göteborg, Sweden.
Division of Chemistry II, Department of Medical Biochemistry and Biophysics, Karolinska Institutet, SE-17 177 Stockholm, Sweden.
J Exp Bot. 2016 Sep;67(17):5133-44. doi: 10.1093/jxb/erw278. Epub 2016 Jul 15.
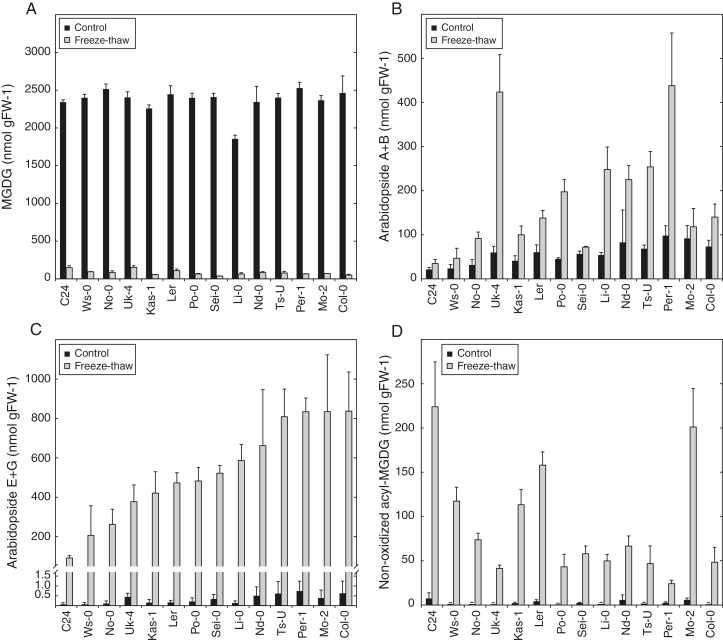
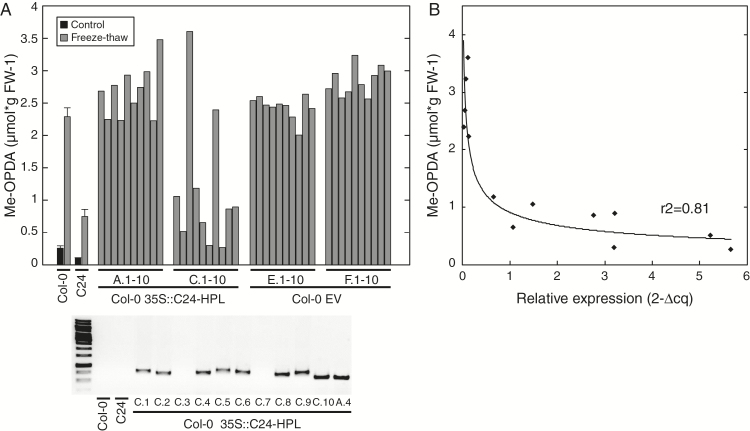
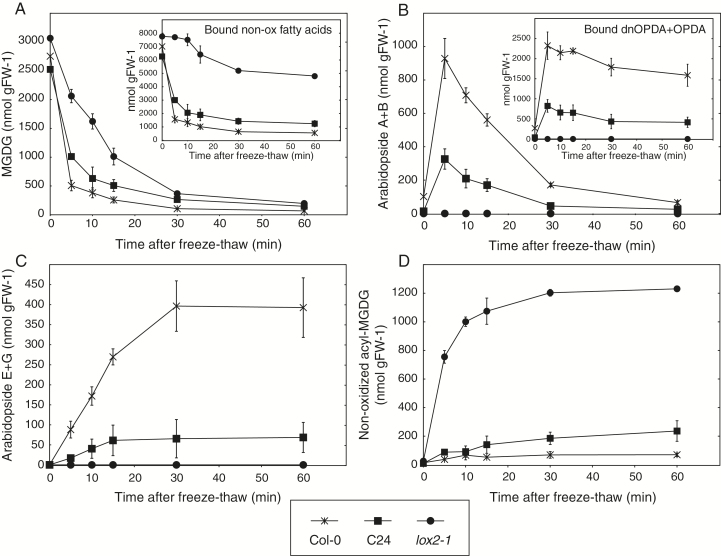
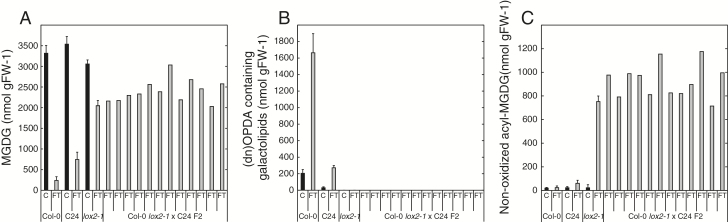
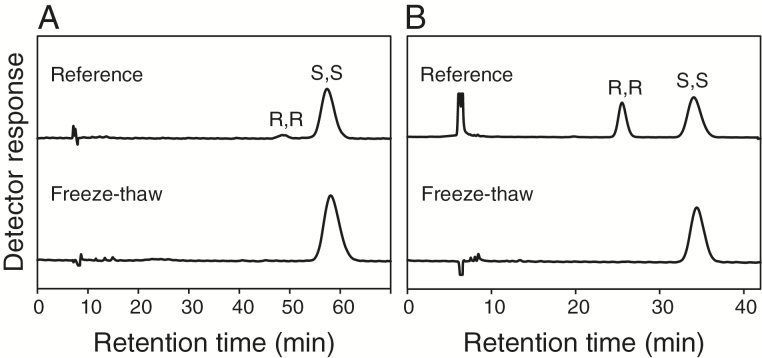
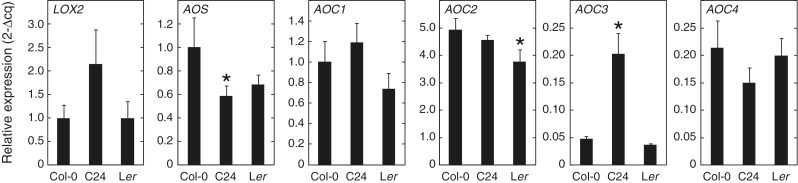
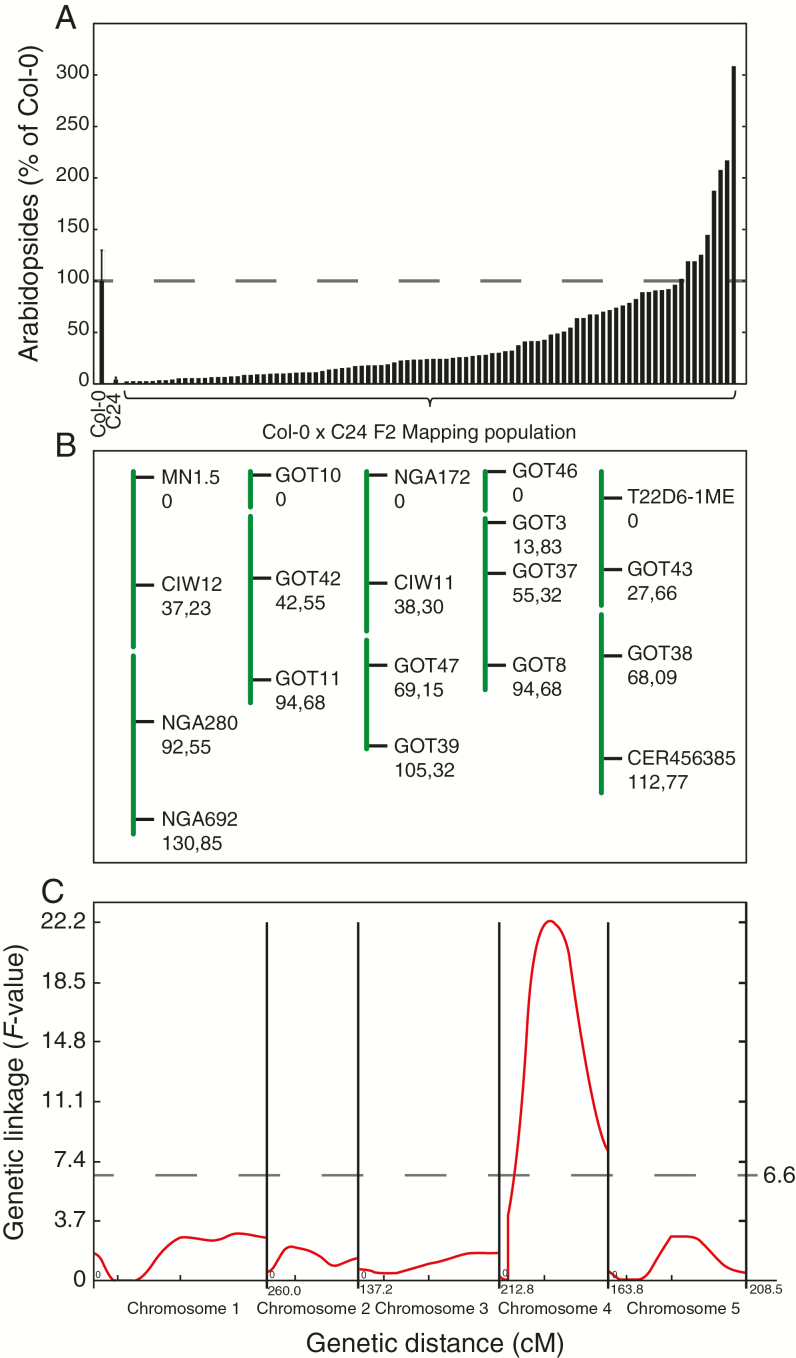
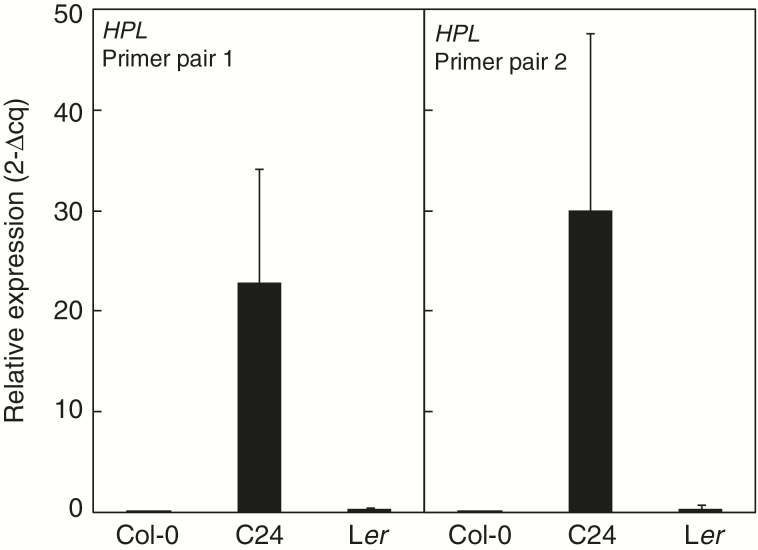
Arabidopsis produces galactolipids containing esters of 12-oxo-phytodienoic acid (OPDA) and dinor-12-oxo-phytodienoic acid (dnOPDA). These lipids are referred to as arabidopsides and accumulate in response to abiotic and biotic stress. We explored the natural genetic variation found in 14 different Arabidopsis accessions to identify genes involved in the formation of arabidopsides. The accession C24 was identified as a poor accumulator of arabidopsides whereas the commonly used accession Col-0 was found to accumulate comparably large amounts of arabidopsides in response to tissue damage. A quantitative trait loci analysis of an F2 population created from a cross between C24 and Col-0 located a region on chromosome four strongly linked to the capacity to form arabidopsides. Expression analysis of HYDROPEROXIDE LYASE 1 (HPL1) showed large differences in transcript abundance between accessions. Transformation of Col-0 plants with the C24 HPL1 allele under transcriptional regulation of the 35S promoter revealed a strong negative correlation between HPL1 expression and arabidopside accumulation after tissue damage, thereby strengthening the view that HPL1 competes with ALLENE OXIDE SYNTHASE (AOS) for lipid-bound hydroperoxide fatty acids. We further show that the last step in the synthesis of galactolipid-bound OPDA and dnOPDA from unstable allene oxides is exclusively enzyme-catalyzed and not the result of spontaneous cyclization. Thus, the results presented here together with previous studies suggest that all steps in arabidopside biosynthesis are enzyme-dependent and apparently all reactions can take place with substrates being esterified to galactolipids.
拟南芥会产生含有12-氧代植物二烯酸(OPDA)和二降-12-氧代植物二烯酸(dnOPDA)酯的半乳糖脂。这些脂质被称为拟南芥苷,并在非生物和生物胁迫下积累。我们探究了14种不同拟南芥生态型中存在的自然遗传变异,以鉴定参与拟南芥苷形成的基因。生态型C24被鉴定为拟南芥苷的低积累型,而常用的生态型Col-0在受到组织损伤时会积累相对大量的拟南芥苷。对由C24和Col-0杂交产生的F2群体进行数量性状基因座分析,确定了四号染色体上的一个区域与形成拟南芥苷的能力紧密相关。对过氧化氢裂解酶1(HPL1)的表达分析表明,不同生态型之间转录本丰度存在很大差异。在35S启动子的转录调控下,用C24 HPL1等位基因转化Col-0植物,结果显示组织损伤后HPL1表达与拟南芥苷积累之间存在强烈的负相关,从而强化了这样一种观点,即HPL1与丙二烯氧化物合酶(AOS)竞争脂质结合的氢过氧化物脂肪酸。我们进一步表明,从不稳定的丙二烯氧化物合成半乳糖脂结合的OPDA和dnOPDA的最后一步完全是由酶催化的,而不是自发环化的结果。因此,本文呈现的结果与先前的研究共同表明,拟南芥苷生物合成的所有步骤都依赖于酶,并且显然所有反应都可以在底物被酯化到半乳糖脂的情况下发生。