Bernal Sierra Yinth A, Haseleu Julia, Kozlenkov Alexey, Bégay Valérie, Lewin Gary R
Department of Neuroscience, Max Delbrück Center for Molecular Medicine in the Helmholtz Association and Charité - Universitätsmedizin Berlin Berlin, Germany.
Front Mol Neurosci. 2017 Mar 15;10:70. doi: 10.3389/fnmol.2017.00070. eCollection 2017.
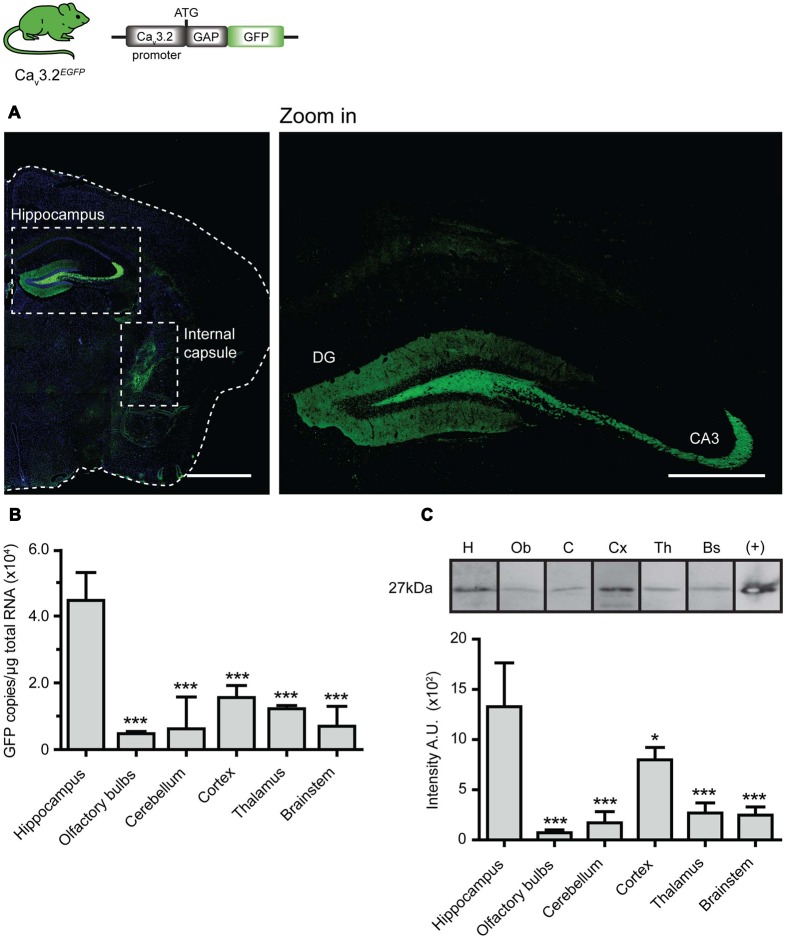
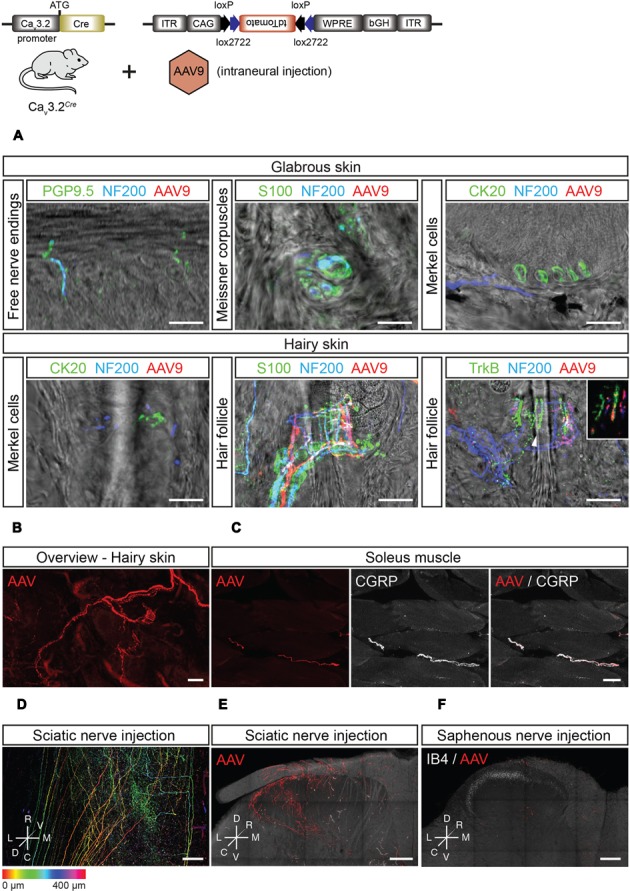
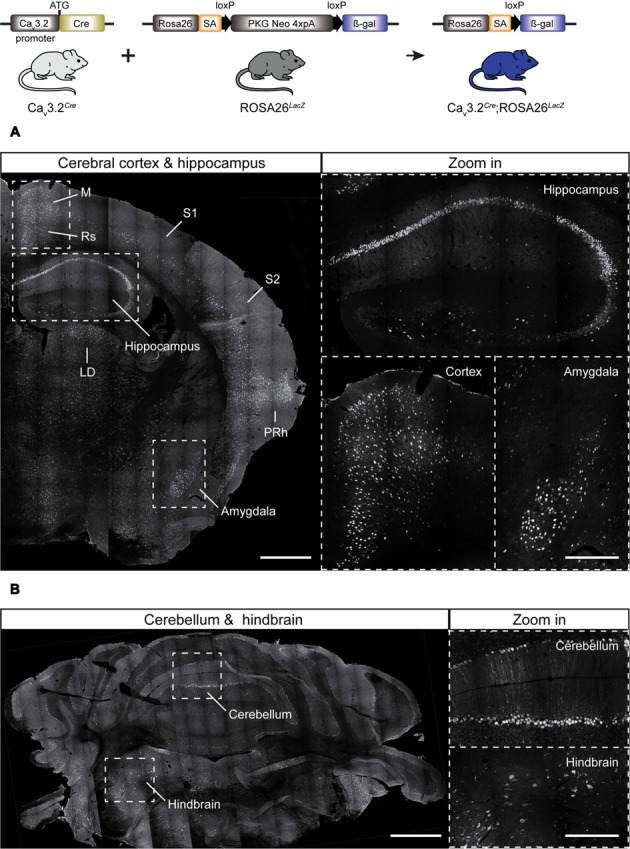
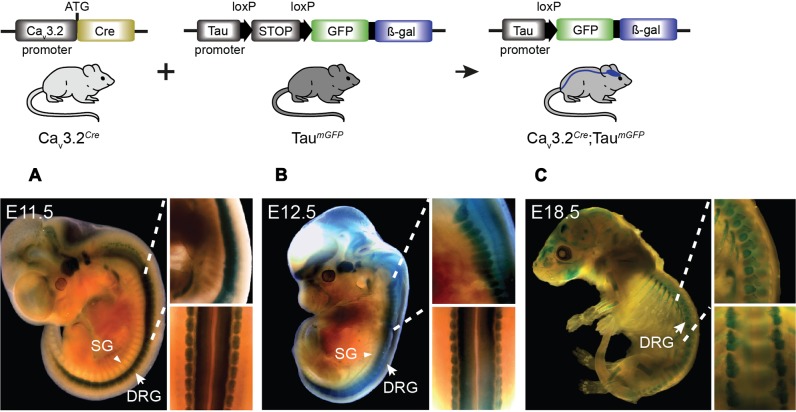
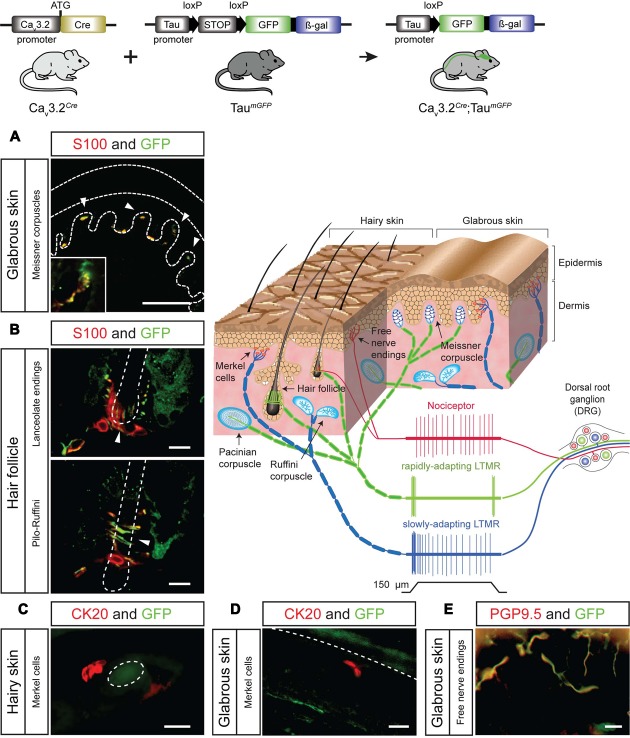
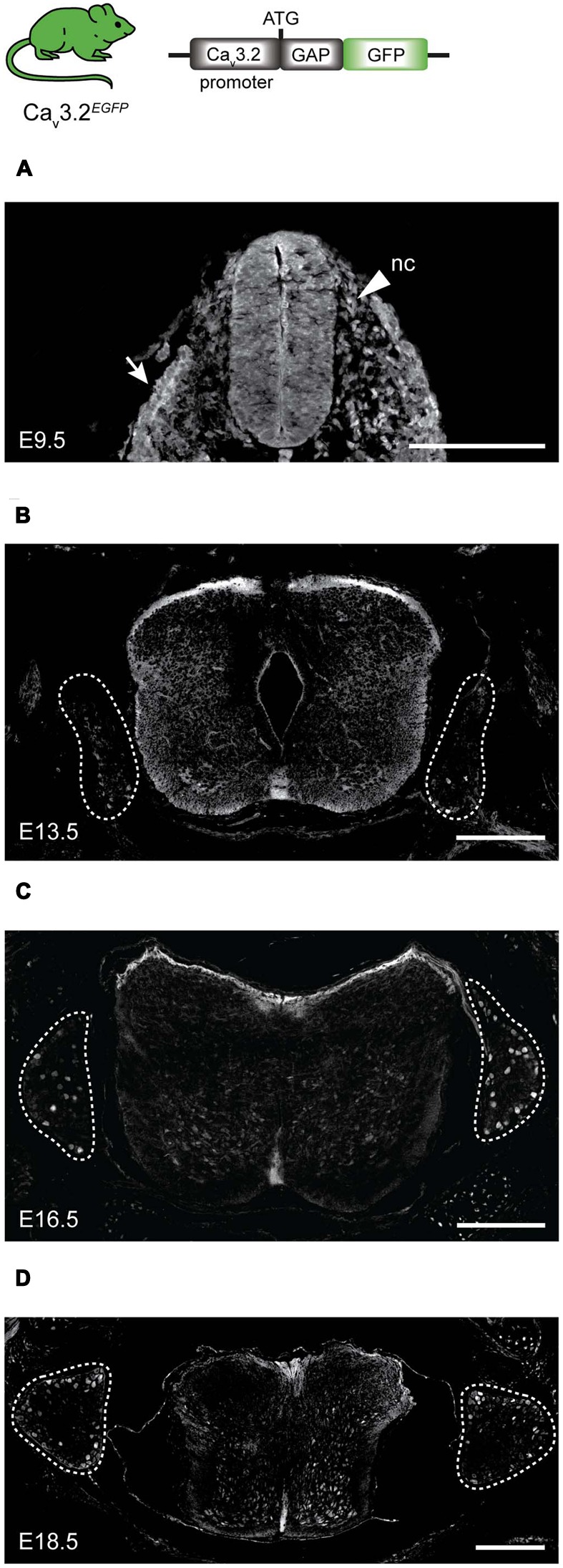
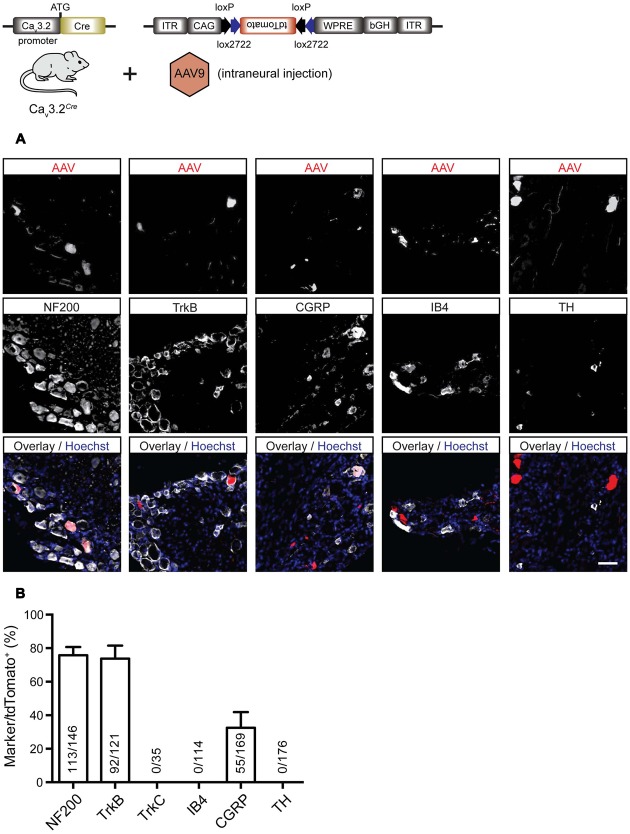
Characterizing the distinct functions of the T-type ion channel subunits Ca3.1, 3.2 or 3.3 has proven difficult due to their highly conserved amino-acid sequences and the lack of pharmacological blockers specific for each subunit. To precisely determine the expression pattern of the Ca3.2 channel in the nervous system we generated two knock-in mouse strains that express EGFP or Cre recombinase under the control of the Ca3.2 gene promoter. We show that in the brains of these animals, the Ca3.2 channel is predominantly expressed in the dentate gyrus of the hippocampus. In the peripheral nervous system, the activation of the promoter starts at E9.5 in neural crest cells that will give rise to dorsal root ganglia (DRG) neurons, but not sympathetic neurons. As development progresses the number of DRG cells expressing the Ca3.2 channel reaches around 7% of the DRG at E16.5, and remains constant until E18.5. Characterization of sensory neuron subpopulations at E18.5 showed that EGFP cells are a heterogeneous population consisting mainly of TrkB and TrkC cells, while only a small percentage of DRG cells were TrkA. Genetic tracing of the sensory nerve end-organ innervation of the skin showed that the activity of the Ca3.2 channel promoter in sensory progenitors marks many mechanoreceptor and nociceptor endings, but spares slowly adapting mechanoreceptors with endings associated with Merkel cells. Our genetic analysis reveals for the first time that progenitors that express the Ca3.2 T-type calcium channel, defines a sensory specific lineage that populates a large proportion of the DRG. Using our Ca3.2-Cre mice together with AAV viruses containing a conditional fluorescent reporter (tdTomato) we could also show that Cre expression is largely restricted to two functionally distinct sensory neuron types in the adult ganglia. Ca3.2 positive neurons innervating the skin were found to only form lanceolate endings on hair follicles and are probably identical to D-hair receptors. A second population of nociceptive sensory neurons expressing the Ca3.2 gene was found to be positive for the calcitonin-gene related peptide but these neurons are deep tissue nociceptors that do not innervate the skin.
由于T型离子通道亚基Ca3.1、3.2和3.3的氨基酸序列高度保守,且缺乏针对每个亚基的特异性药理阻断剂,因此很难确定它们各自独特的功能。为了精确确定Ca3.2通道在神经系统中的表达模式,我们构建了两种基因敲入小鼠品系,它们在Ca3.2基因启动子的控制下表达EGFP或Cre重组酶。我们发现,在这些动物的大脑中,Ca3.2通道主要表达于海马齿状回。在周围神经系统中,启动子的激活在E9.5时开始于神经嵴细胞,这些细胞将发育为背根神经节(DRG)神经元,但不包括交感神经元。随着发育进程,在E16.5时表达Ca3.2通道的DRG细胞数量达到DRG总数的7%左右,并在E18.5之前保持稳定。对E18.5时感觉神经元亚群的特征分析表明,EGFP阳性细胞是一个异质性群体,主要由TrkB和TrkC细胞组成,而只有一小部分DRG细胞是TrkA阳性。对皮肤感觉神经终末器官支配的基因追踪显示,感觉祖细胞中Ca3.2通道启动子的活性标记了许多机械感受器和伤害感受器的终末,但不包括与默克尔细胞相关的慢适应性机械感受器的终末。我们的基因分析首次揭示,表达Ca3.2 T型钙通道的祖细胞定义了一个感觉特异性谱系,该谱系占据了大部分DRG。使用我们的Ca3.2-Cre小鼠以及含有条件荧光报告基因(tdTomato)的腺相关病毒(AAV),我们还发现,在成年神经节中,Cre表达主要局限于两种功能不同的感觉神经元类型。支配皮肤的Ca3.2阳性神经元仅在毛囊上形成柳叶状终末,可能与D型毛感受器相同。另一群表达Ca3.2基因的伤害性感觉神经元对降钙素基因相关肽呈阳性,但这些神经元是深部组织伤害感受器,不支配皮肤。