Wang Chien-Hsiang, Huang Yi-Chun, Chen Pei-Yi, Cheng Ying-Ju, Kao Hsiu-Hua, Pi Haiwei, Chien Cheng-Ting
Institute of Molecular Biology, Academia Sinica, Taipei, Taiwan.
Institute of Neuroscience, National Yang Ming University, Taipei, Taiwan.
Elife. 2017 May 10;6:e26886. doi: 10.7554/eLife.26886.
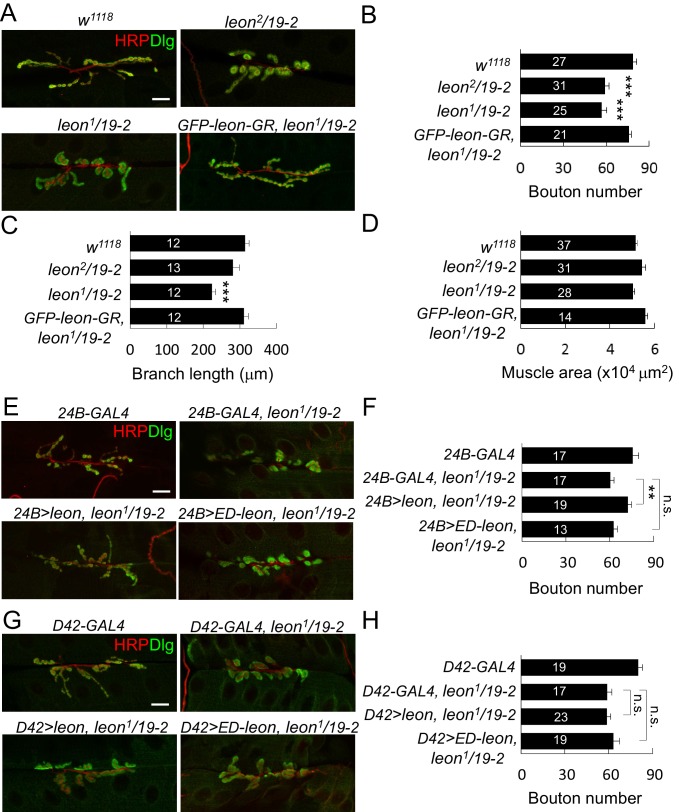
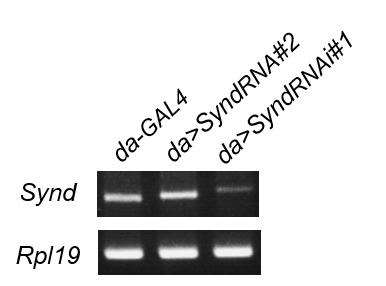
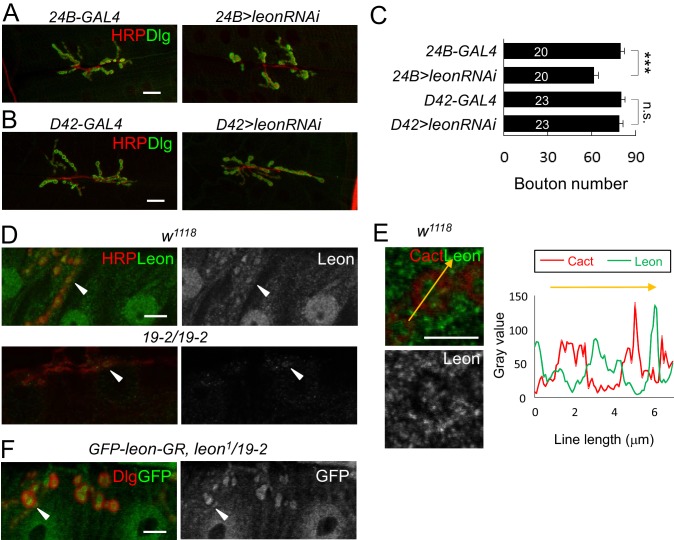
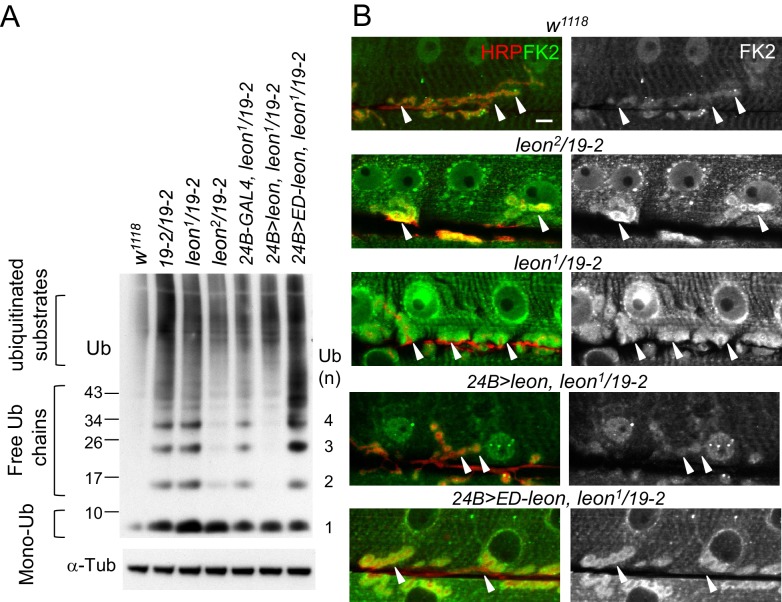
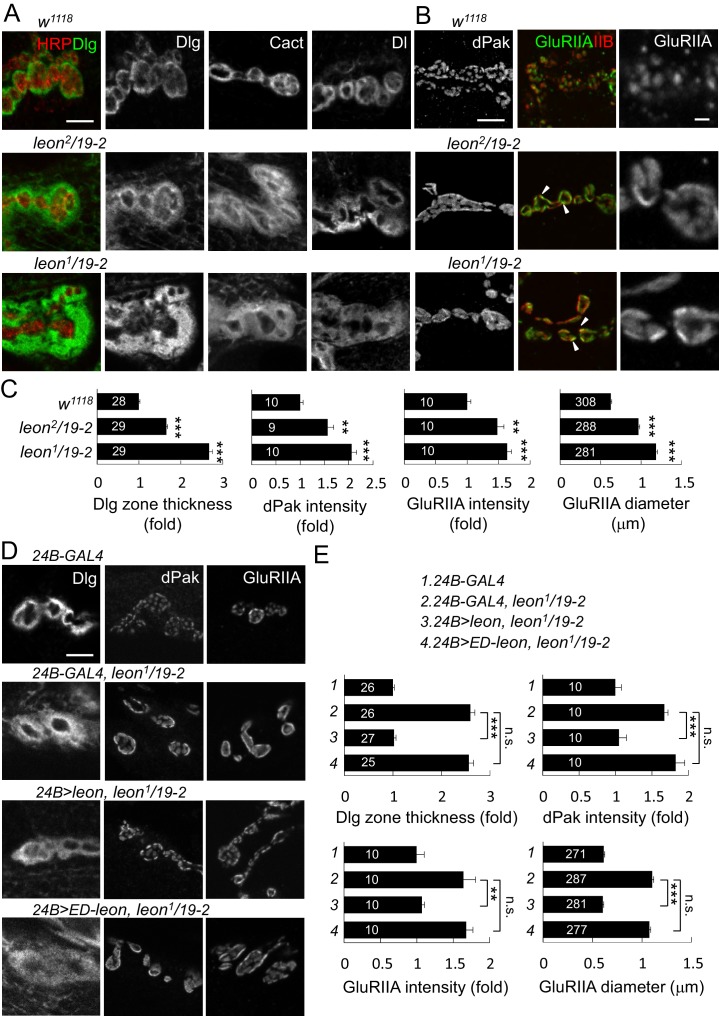
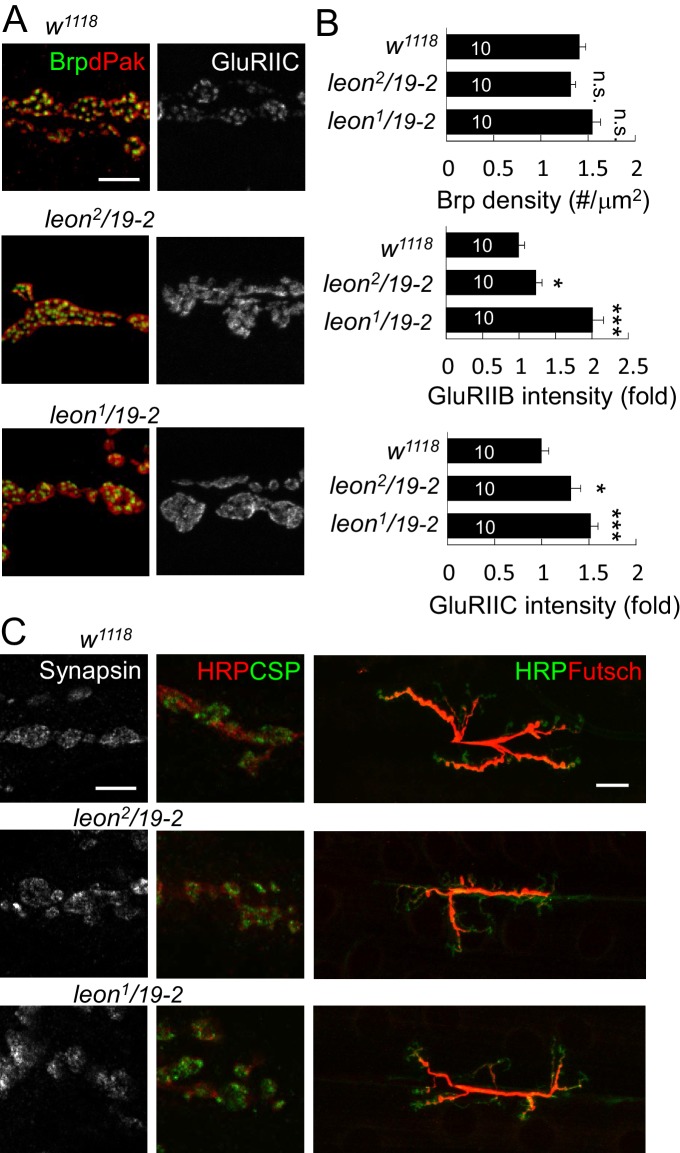
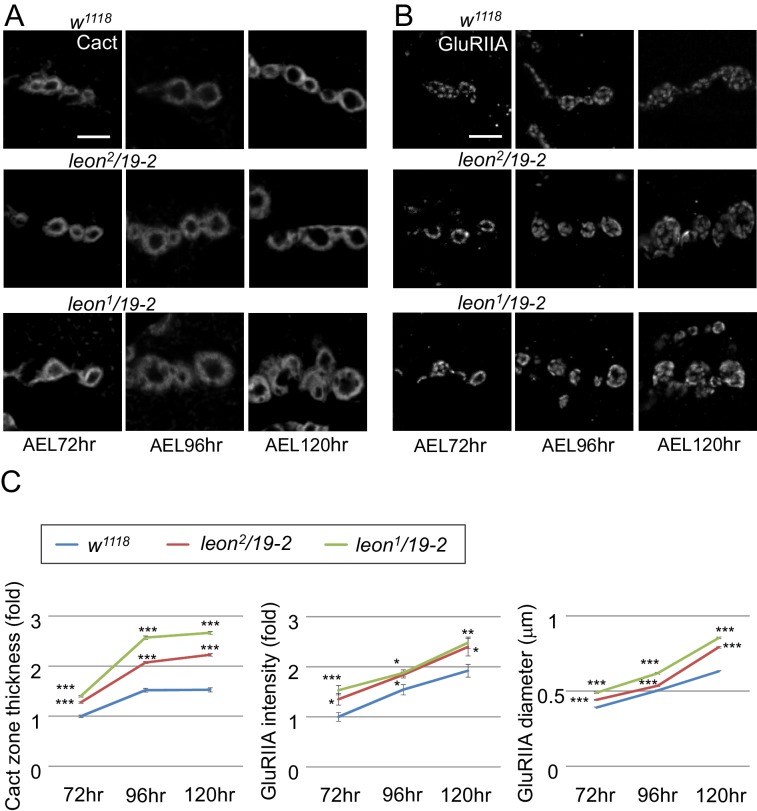
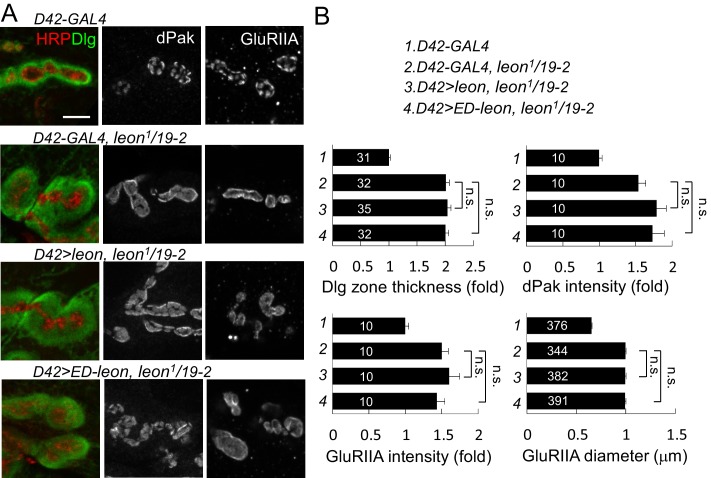
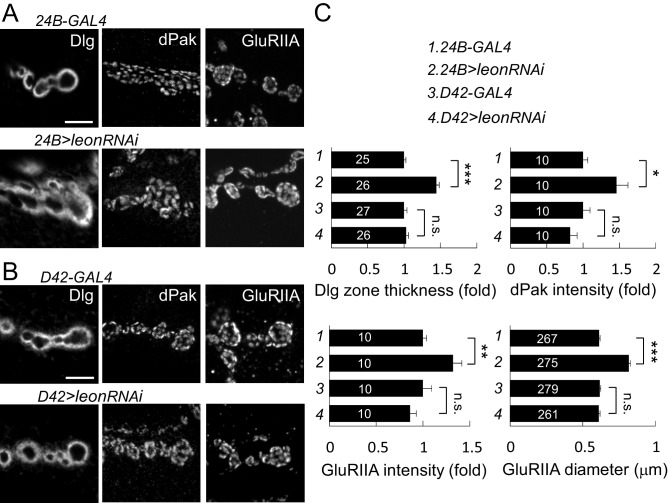
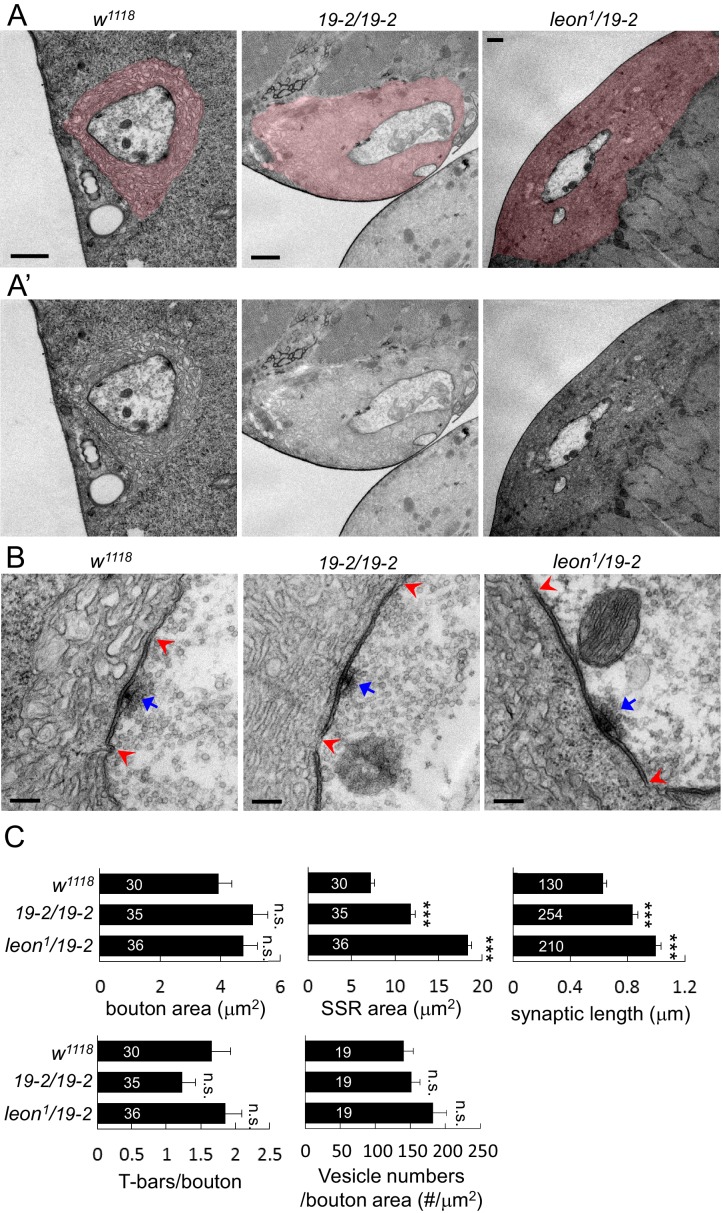
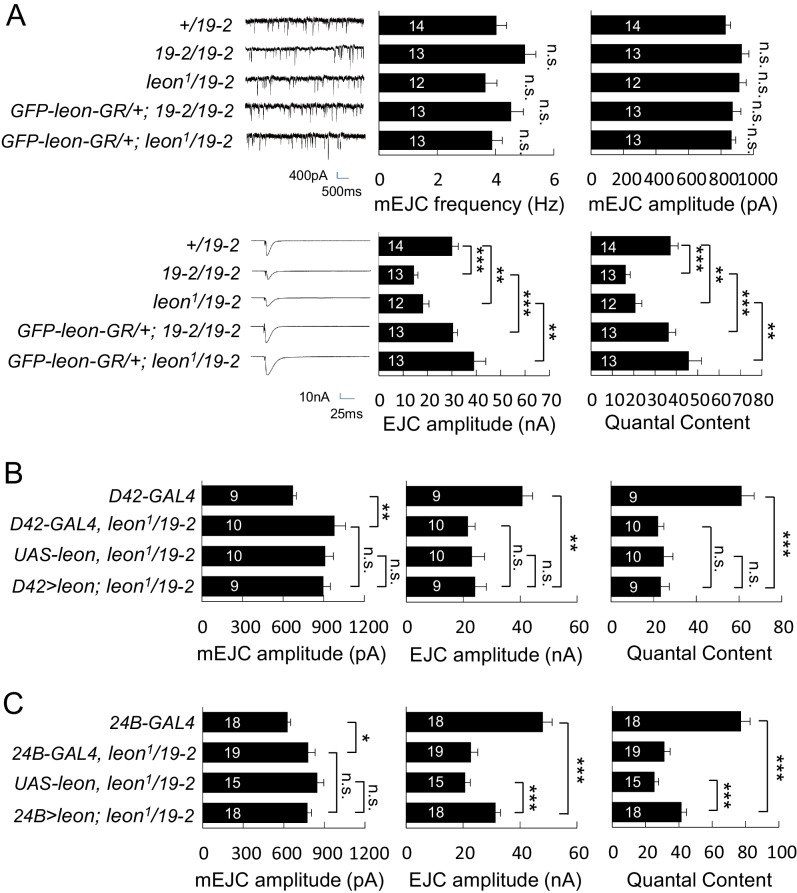
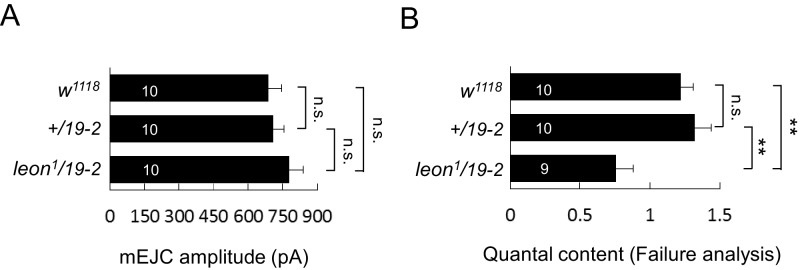
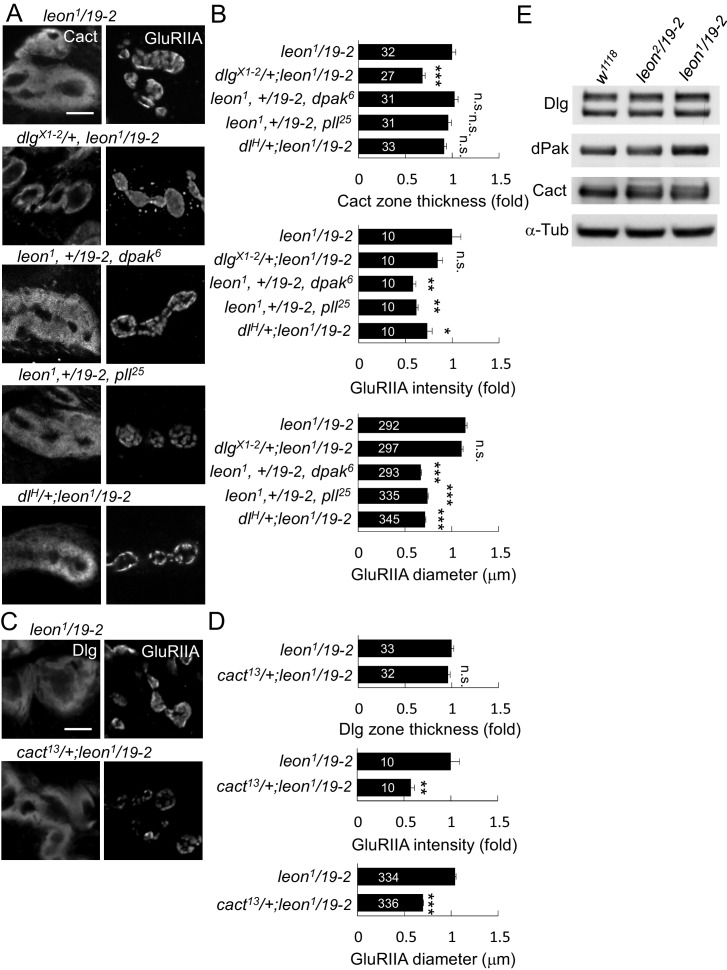
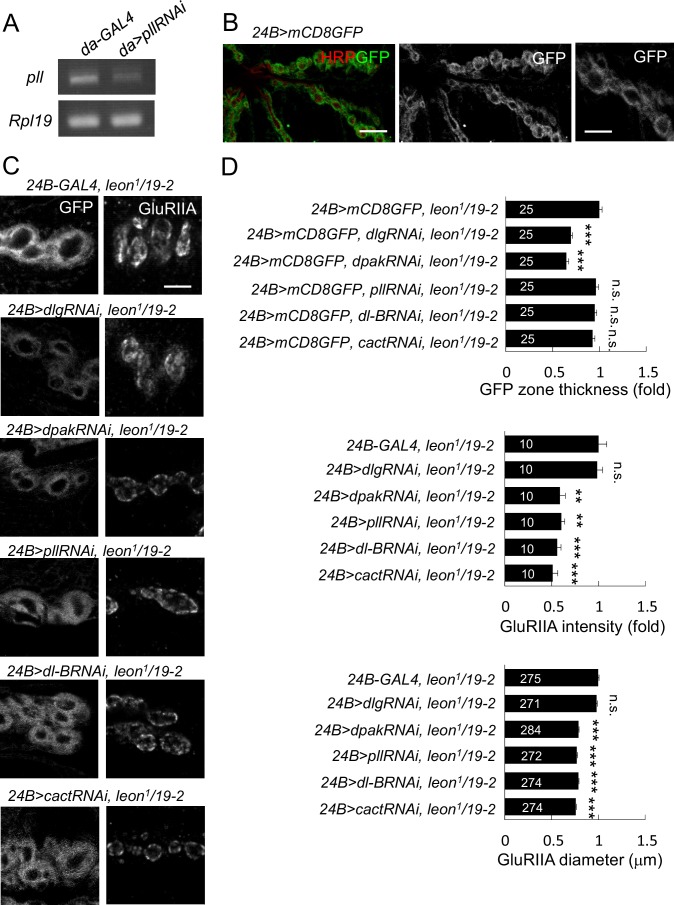
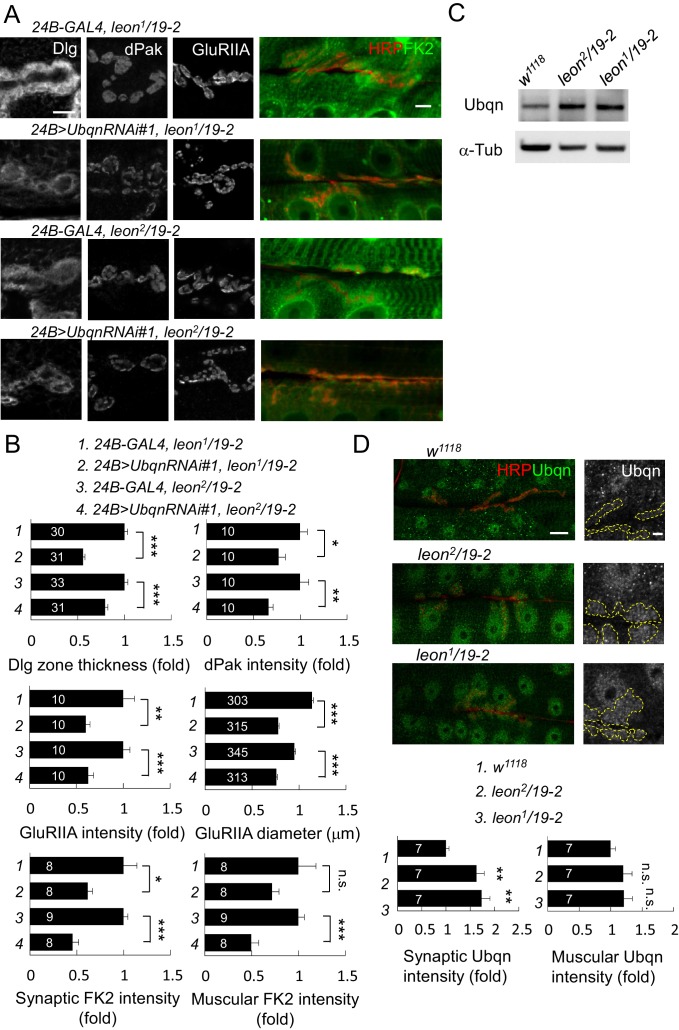
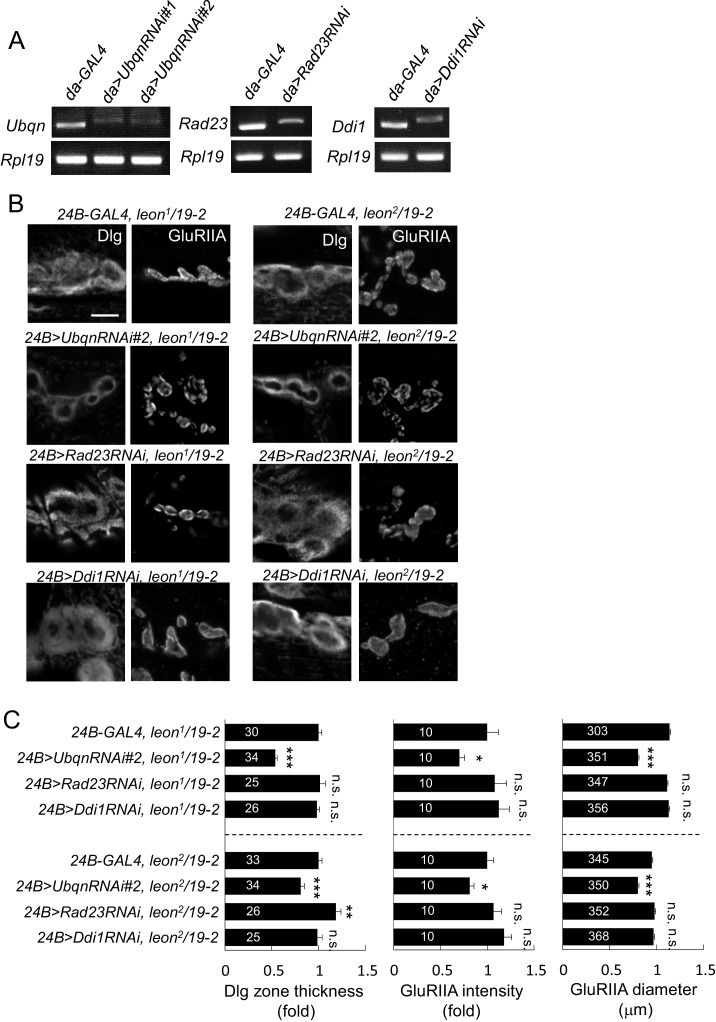
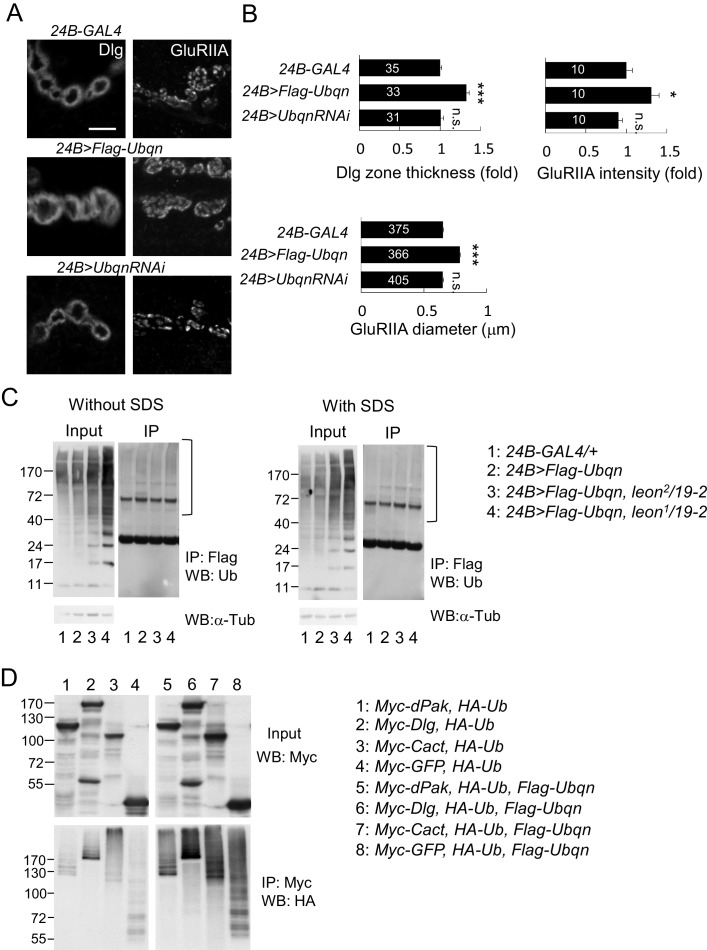
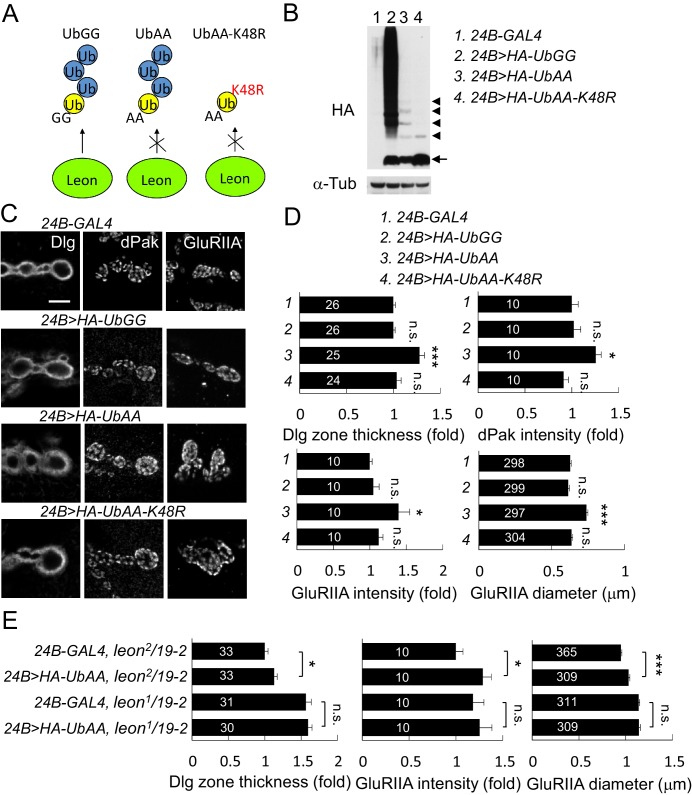
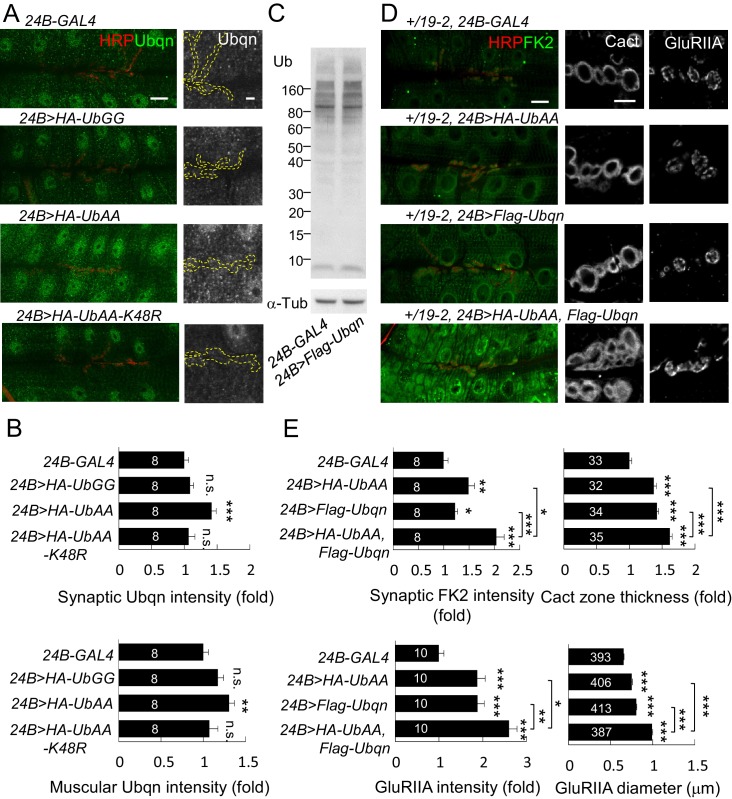
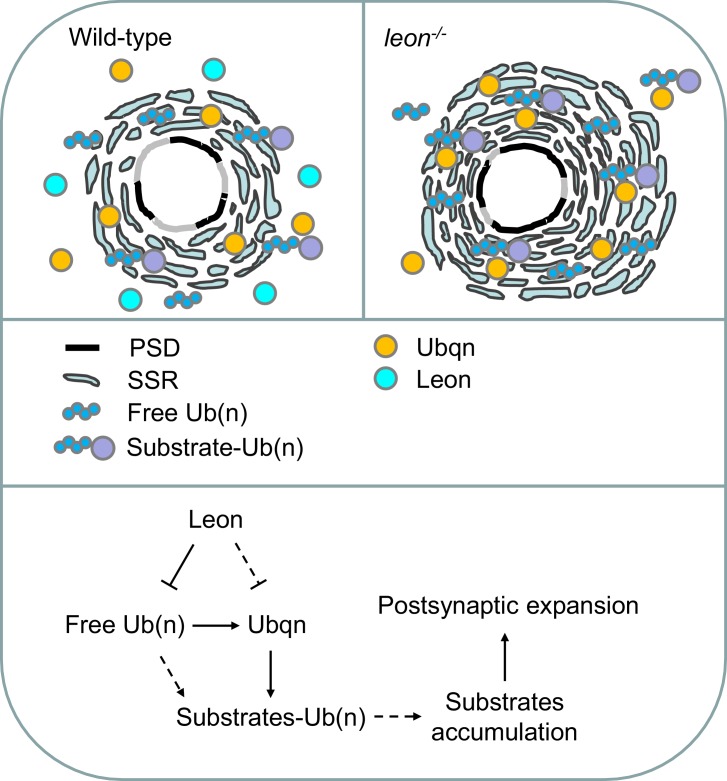
Synapse formation and growth are tightly controlled processes. How synaptic growth is terminated after reaching proper size remains unclear. Here, we show that Leon, the USP5 deubiquitinase, controls postsynaptic growth. In mutants, postsynaptic specializations of neuromuscular junctions are dramatically expanded, including the subsynaptic reticulum, the postsynaptic density, and the glutamate receptor cluster. Expansion of these postsynaptic features is caused by a disruption of ubiquitin homeostasis with accumulation of free ubiquitin chains and ubiquitinated substrates in the mutant. Accumulation of Ubiquilin (Ubqn), the ubiquitin receptor whose human homolog ubiquilin 2 is associated with familial amyotrophic lateral sclerosis, also contributes to defects in postsynaptic growth and ubiquitin homeostasis. Importantly, accumulations of postsynaptic proteins cause different aspects of postsynaptic overgrowth in mutants. Thus, the deubiquitinase Leon maintains ubiquitin homeostasis and proper Ubqn levels, preventing postsynaptic proteins from accumulation to confine postsynaptic growth.
突触的形成和生长是受到严格控制的过程。在达到适当大小后,突触生长是如何终止的仍不清楚。在这里,我们表明,USP5去泛素化酶Leon控制着突触后生长。在突变体中,神经肌肉接头的突触后特化显著扩展,包括突触下网状结构、突触后致密区和谷氨酸受体簇。这些突触后特征的扩展是由泛素稳态的破坏引起的,在突变体中存在游离泛素链和泛素化底物的积累。泛素受体Ubiquilin(Ubqn)的积累也导致突触后生长缺陷和泛素稳态异常,其人类同源物泛素连接蛋白2与家族性肌萎缩侧索硬化症相关。重要的是,突触后蛋白的积累在突变体中导致了突触后过度生长的不同方面。因此,去泛素化酶Leon维持泛素稳态和适当的Ubqn水平,防止突触后蛋白积累以限制突触后生长。