The Black Family Stem Cell Institute, Icahn School of Medicine at Mount Sinai, New York, United States.
Department of Cell, Developmental and Regenerative Biology, Icahn School of Medicine at Mount Sinai, New York, United States.
Elife. 2017 Nov 23;6:e33333. doi: 10.7554/eLife.33333.
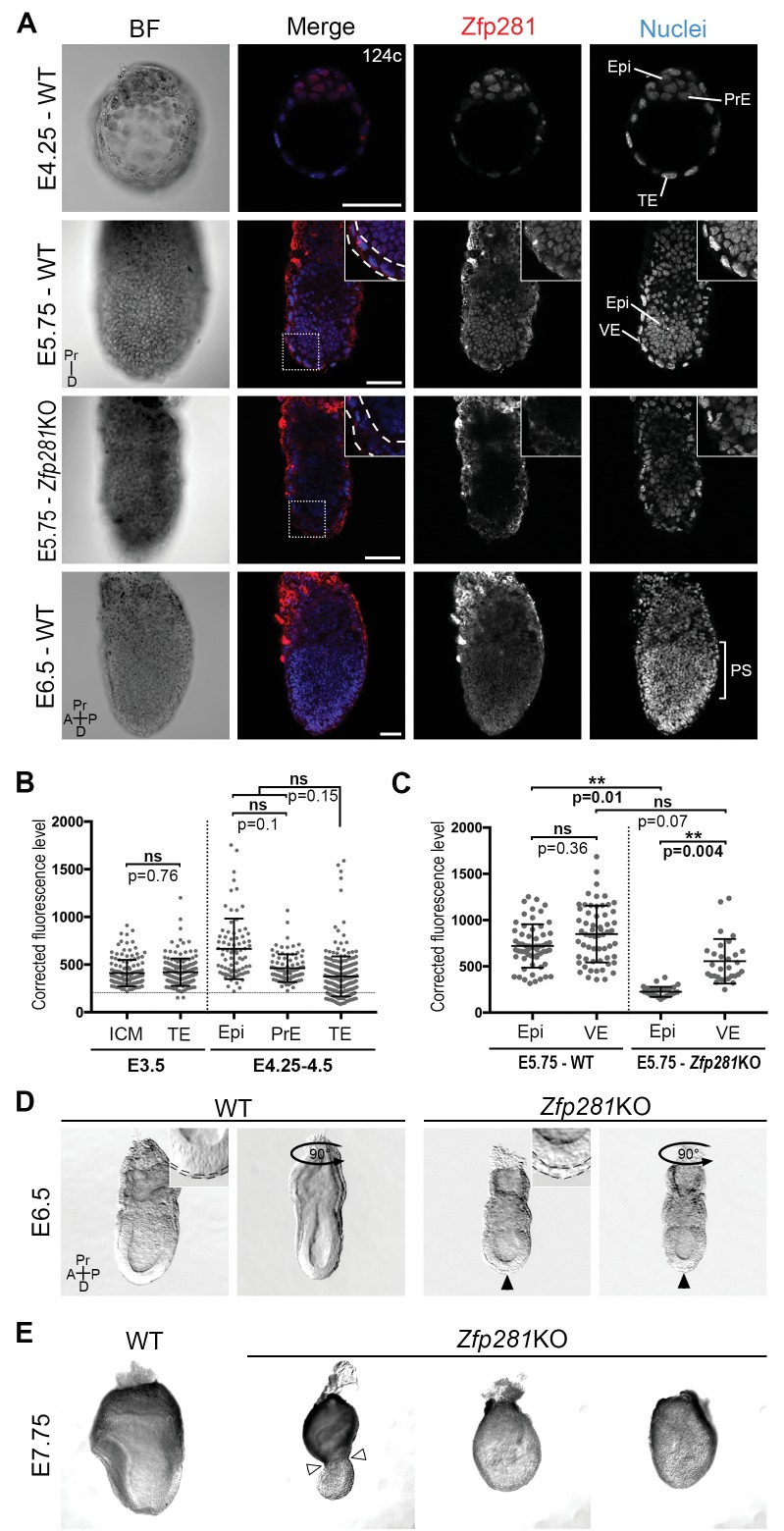
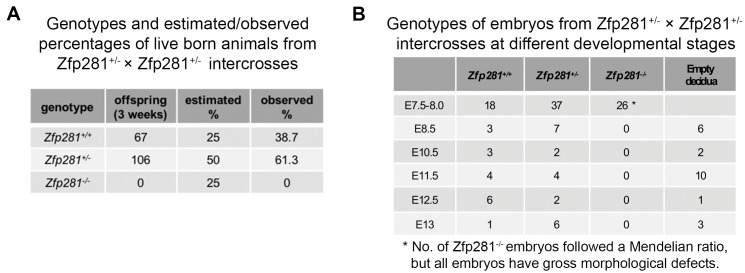
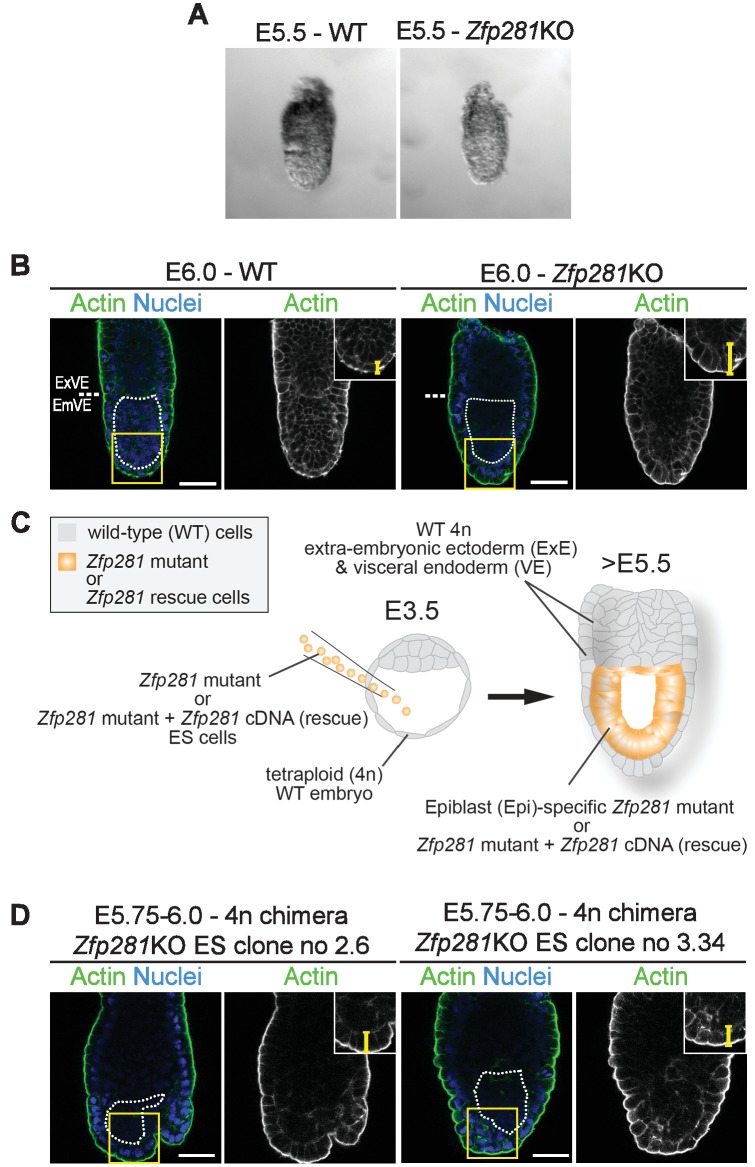
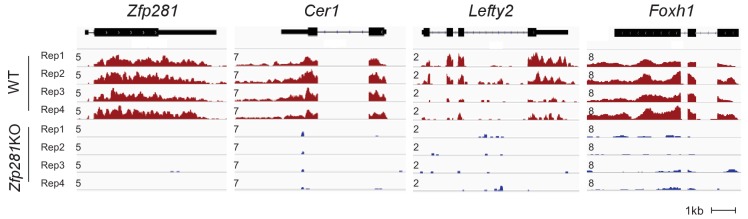
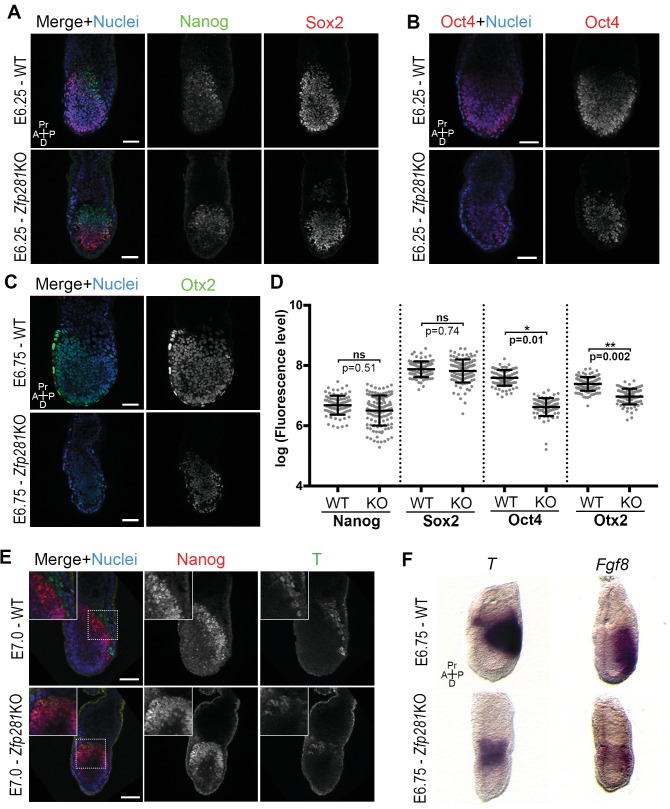
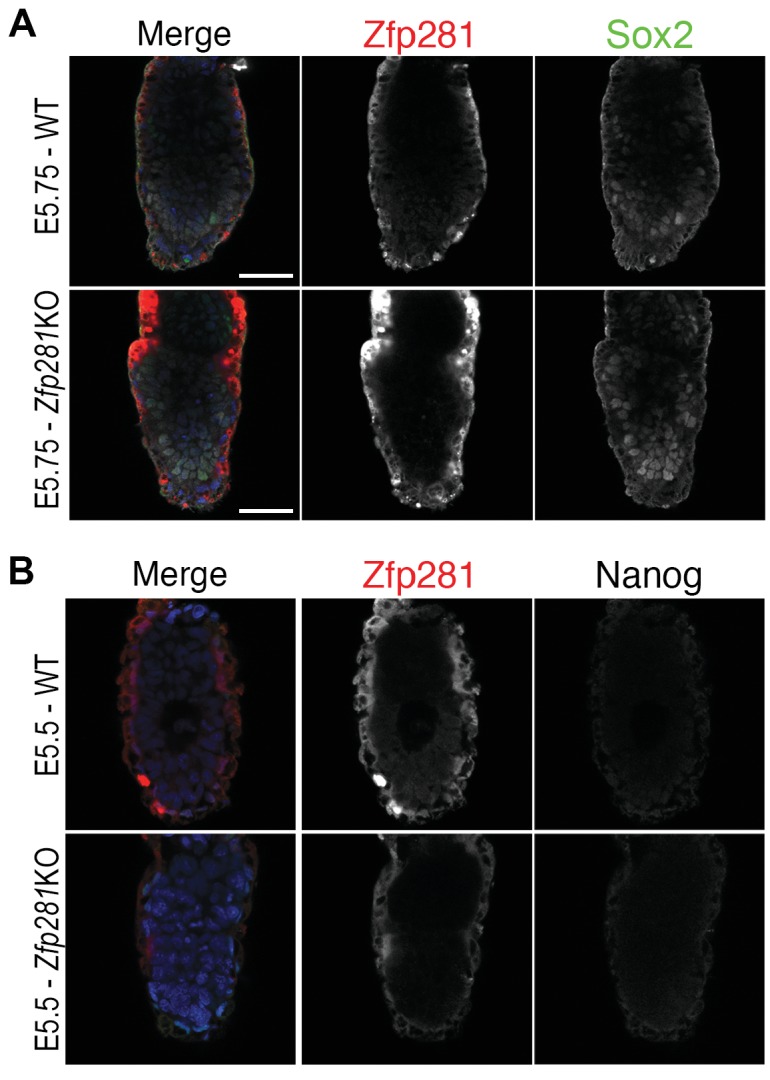
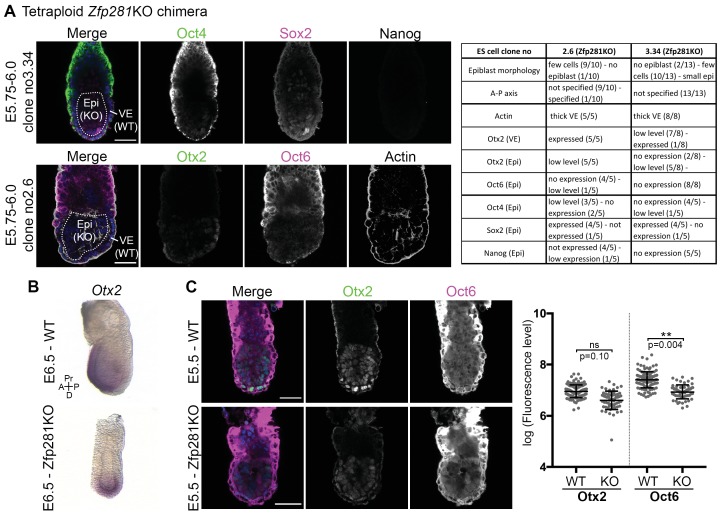
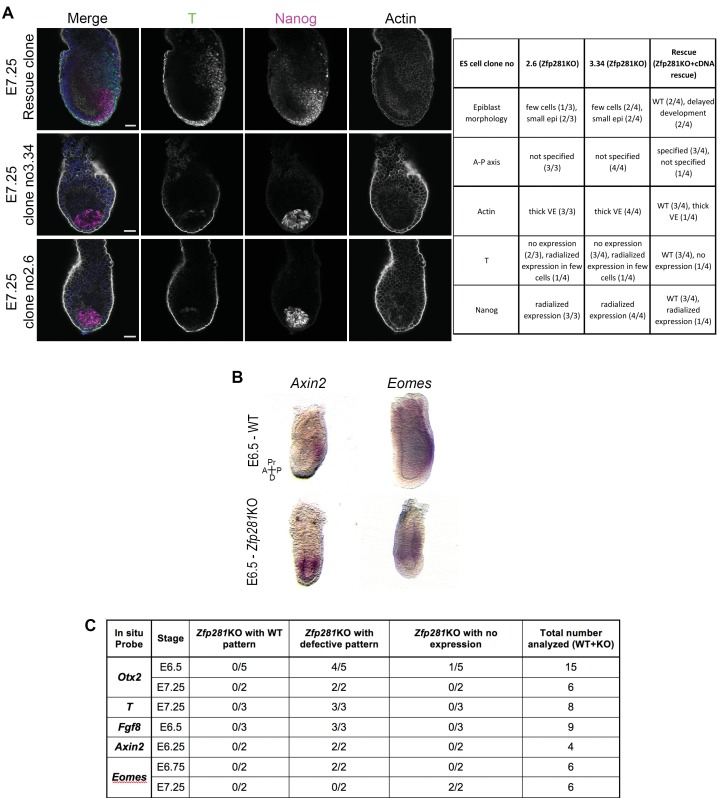
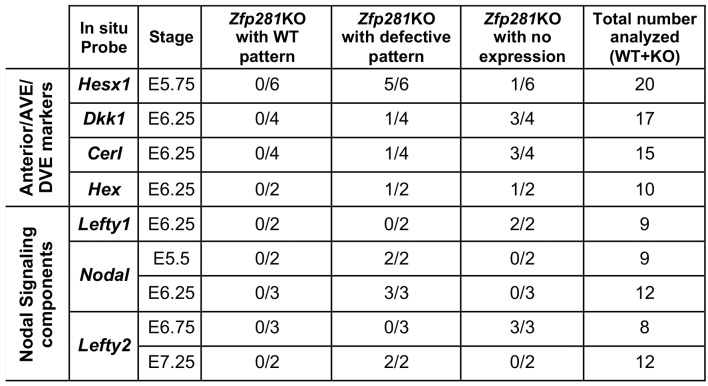
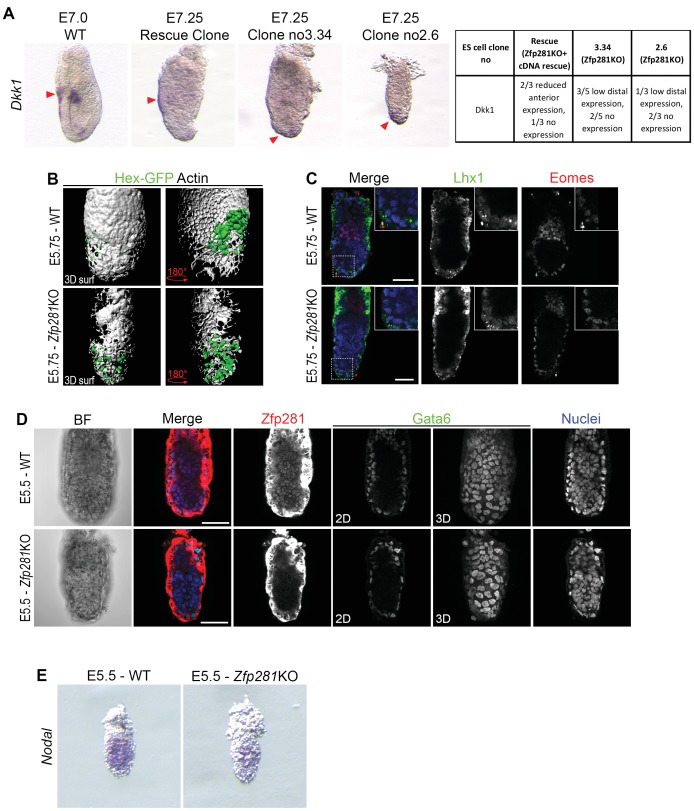
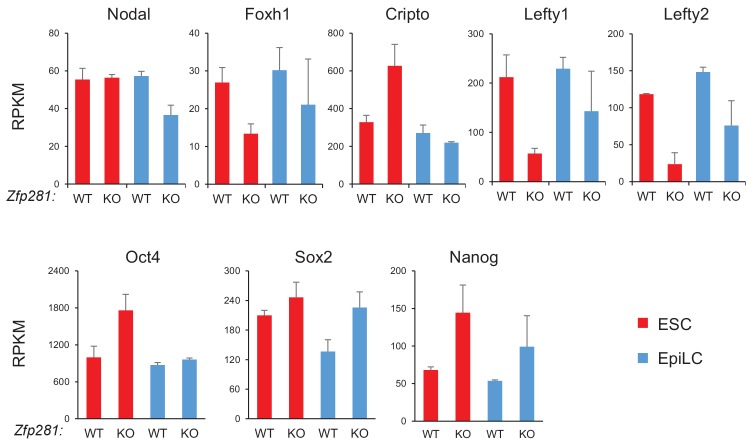
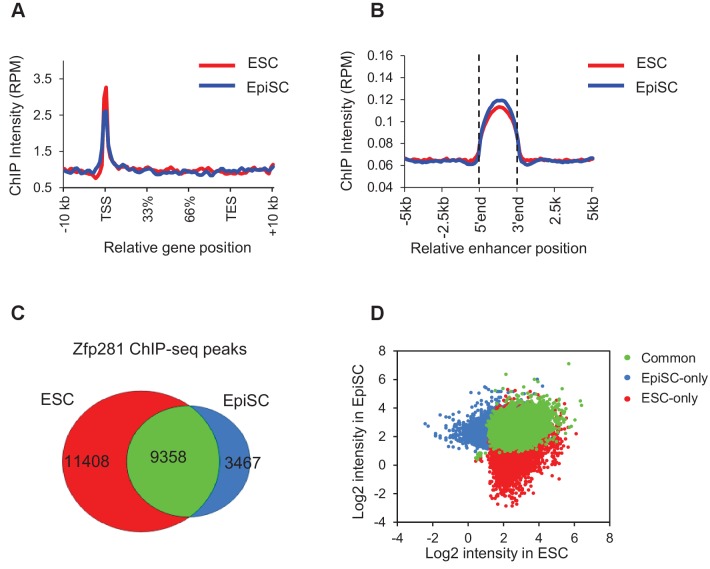
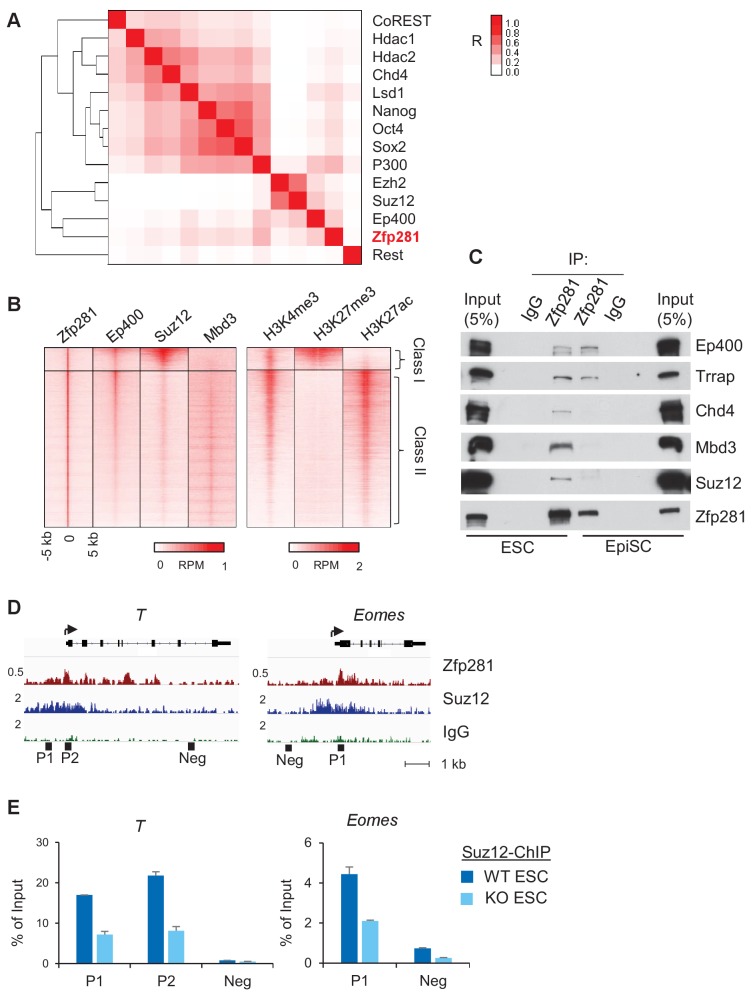
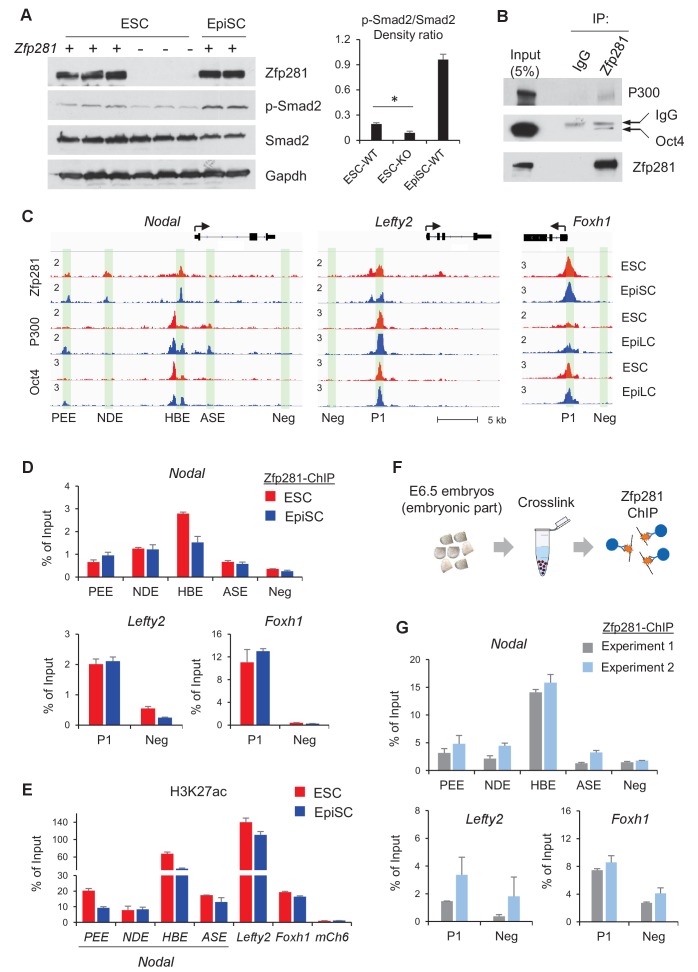
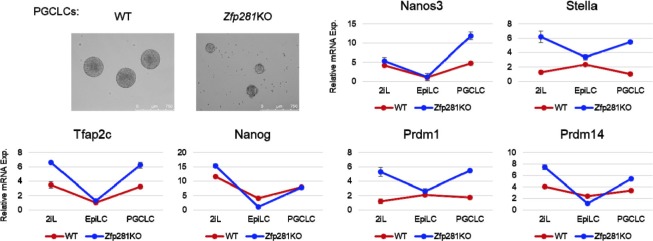
Pluripotency is defined by a cell's potential to differentiate into any somatic cell type. How pluripotency is transited during embryo implantation, followed by cell lineage specification and establishment of the basic body plan, is poorly understood. Here we report the transcription factor Zfp281 functions in the exit from naive pluripotency occurring coincident with pre-to-post-implantation mouse embryonic development. By characterizing mutant phenotypes and identifying Zfp281 gene targets and protein partners in developing embryos and cultured pluripotent stem cells, we establish critical roles for Zfp281 in activating components of the Nodal signaling pathway and lineage-specific genes. Mechanistically, Zfp281 cooperates with histone acetylation and methylation complexes at target gene enhancers and promoters to exert transcriptional activation and repression, as well as epigenetic control of epiblast maturation leading up to anterior-posterior axis specification. Our study provides a comprehensive molecular model for understanding pluripotent state progressions in vivo during mammalian embryonic development.
多能性是由细胞分化为任何体细胞类型的潜力所定义的。在胚胎植入过程中,多能性是如何过渡的,随后是细胞谱系特化和基本身体计划的建立,这一点还不太清楚。在这里,我们报告转录因子 Zfp281 在与小鼠胚胎发育的预着床到着床后同时发生的原始多能性退出中发挥作用。通过描述突变表型,并鉴定发育中的胚胎和成体多能干细胞中 Zfp281 的基因靶标和蛋白伴侣,我们确定 Zfp281 在激活 Nodal 信号通路和谱系特异性基因的组成部分中具有关键作用。从机制上讲,Zfp281 在靶基因增强子和启动子处与组蛋白乙酰化和甲基化复合物合作,发挥转录激活和抑制作用,以及对前后轴特化前的内胚层成熟的表观遗传控制。我们的研究为理解哺乳动物胚胎发育过程中体内多能状态的进展提供了一个全面的分子模型。