Department of Infection, Immunity, and Inflammation, University of Leicester, Leicester LE1 9HN, United Kingdom.
Leicester Institute of Structural and Chemical Biology, University of Leicester, Leicester LE1 7RH, United Kingdom.
Proc Natl Acad Sci U S A. 2018 Jan 23;115(4):768-773. doi: 10.1073/pnas.1718709115. Epub 2018 Jan 8.
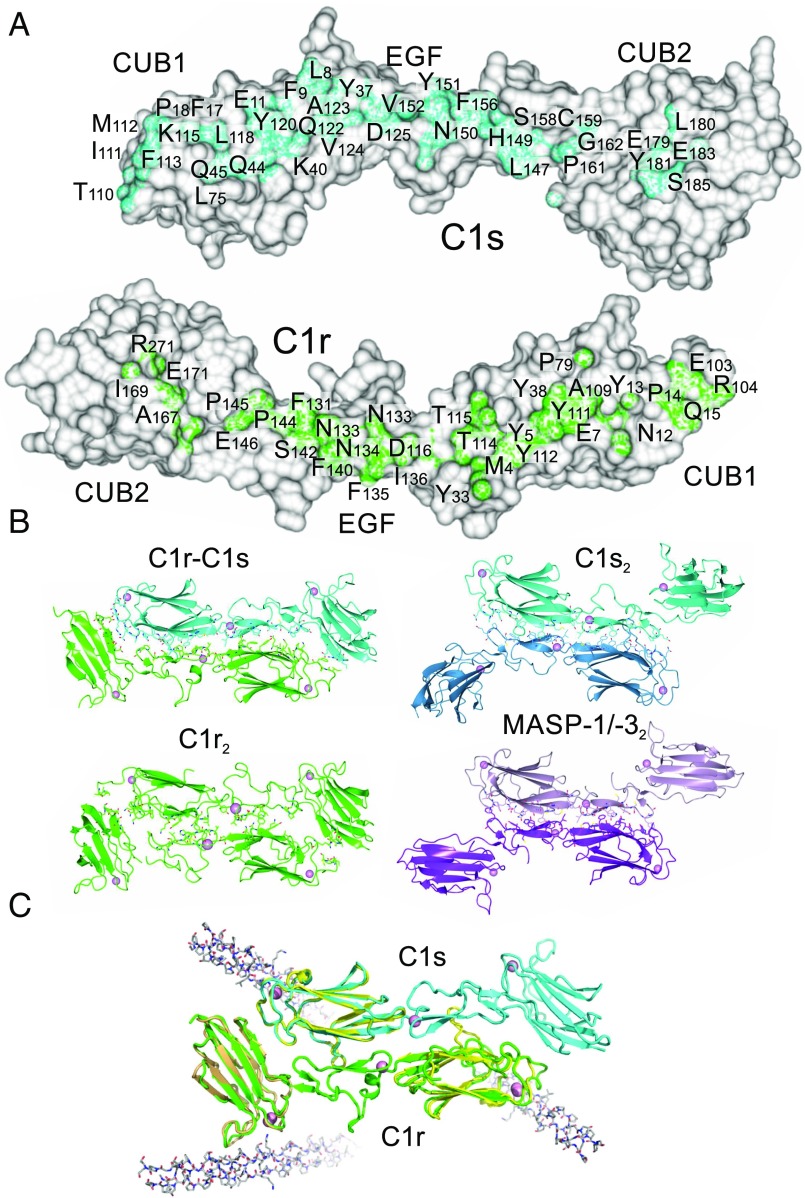
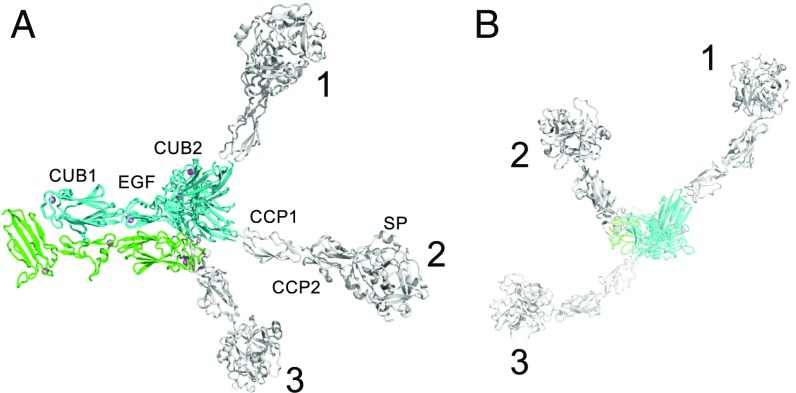
The multiprotein complex C1 initiates the classical pathway of complement activation on binding to antibody-antigen complexes, pathogen surfaces, apoptotic cells, and polyanionic structures. It is formed from the recognition subcomponent C1q and a tetramer of proteases C1rC1s as a Ca-dependent complex. Here we have determined the structure of a complex between the CUB1-EGF-CUB2 fragments of C1r and C1s to reveal the C1r-C1s interaction that forms the core of C1. Both fragments are L-shaped and interlock to form a compact antiparallel heterodimer with a Ca from each subcomponent at the interface. Contacts, involving all three domains of each protease, are more extensive than those of C1r or C1s homodimers, explaining why heterocomplexes form preferentially. The available structural and biophysical data support a model of C1rC1s in which two C1r-C1s dimers are linked via the catalytic domains of C1r. They are incompatible with a recent model in which the N-terminal domains of C1r and C1s form a fixed tetramer. On binding to C1q, the proteases become more compact, with the C1r-C1s dimers at the center and the six collagenous stems of C1q arranged around the perimeter. Activation is likely driven by separation of the C1r-C1s dimer pairs when C1q binds to a surface. Considerable flexibility in C1s likely facilitates C1 complex formation, activation of C1s by C1r, and binding and activation of downstream substrates C4 and C4b-bound C2 to initiate the reaction cascade.
多蛋白复合物 C1 与抗体-抗原复合物、病原体表面、凋亡细胞和多阴离子结构结合后,可启动补体经典途径的激活。它由识别亚基 C1q 和作为 Ca 依赖性复合物的蛋白酶 C1rC1s 四聚体组成。在此,我们确定了 C1r 和 C1s 的 CUB1-EGF-CUB2 片段之间复合物的结构,以揭示形成 C1 核心的 C1r-C1s 相互作用。这两个片段都是 L 形的,相互交错形成一个紧凑的反平行异二聚体,每个亚基的 Ca 位于界面处。涉及每个蛋白酶的三个结构域的接触比 C1r 或 C1s 同源二聚体的接触更广泛,这解释了为什么异源复合物优先形成。现有的结构和生物物理数据支持 C1rC1s 的模型,其中两个 C1r-C1s 二聚体通过 C1r 的催化结构域连接。该模型与最近的模型不一致,后者认为 C1r 和 C1s 的 N 端结构域形成固定的四聚体。与 C1q 结合后,蛋白酶变得更加紧凑,C1r-C1s 二聚体位于中心,C1q 的六个胶原样茎围绕周边排列。当 C1q 与表面结合时,C1r-C1s 二聚体的分离可能驱动激活。C1s 的相当大的灵活性可能有助于 C1 复合物的形成、C1r 对 C1s 的激活以及下游底物 C4 和 C4b 结合的 C2 的结合和激活,从而启动反应级联。