Department of Chemistry, Graduate School of Science, Shizuoka University, 836 Ohya, Suruga-ku, Shizuoka, 422-8529, Japan.
Research Institute of Green Science and Technology, Shizuoka University, 836 Ohya, Suruga-ku, Shizuoka, 422-8529, Japan.
Nucleic Acids Res. 2018 Jul 6;46(12):5894-5901. doi: 10.1093/nar/gky391.
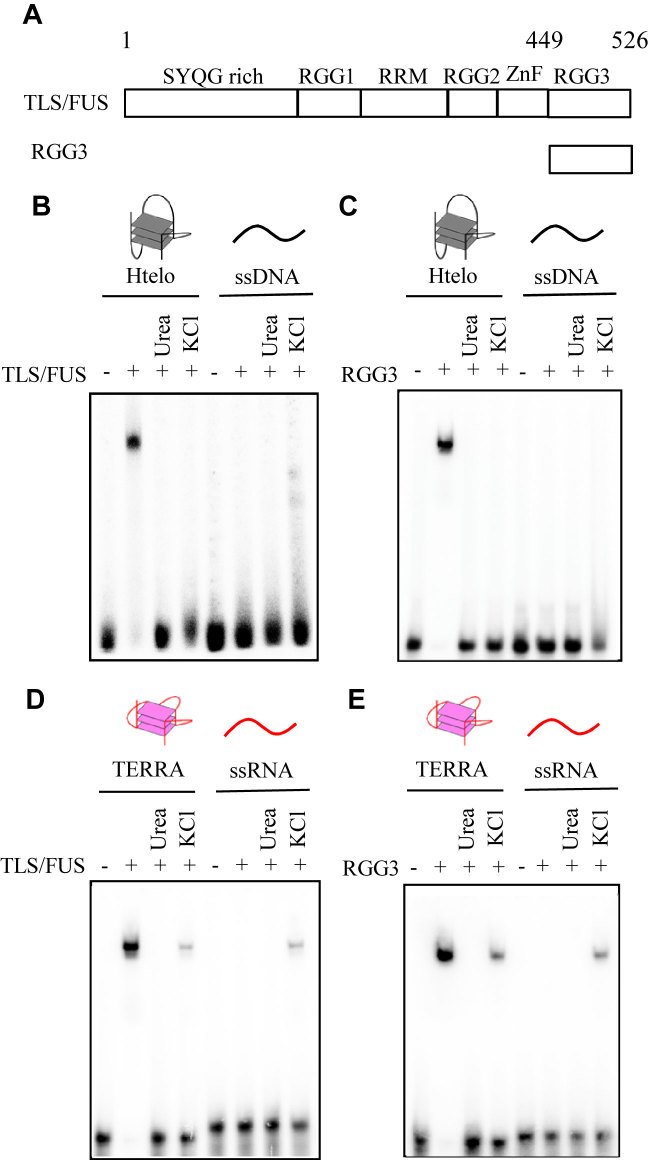
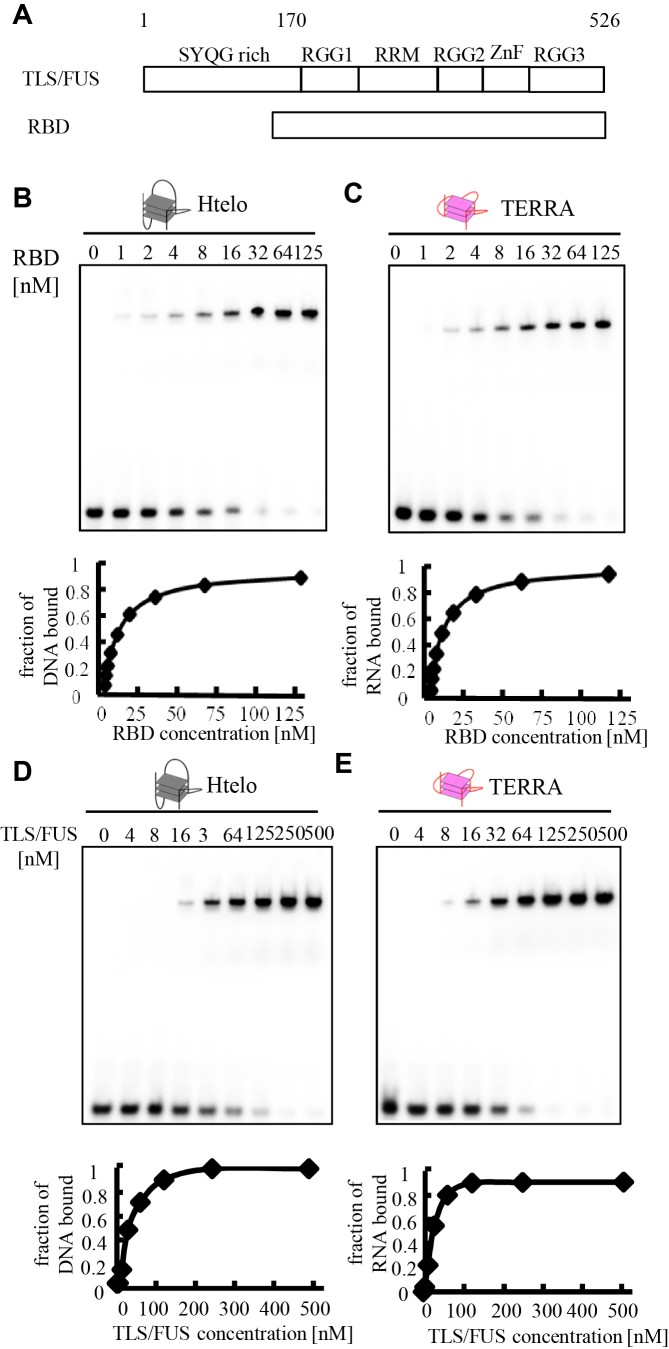
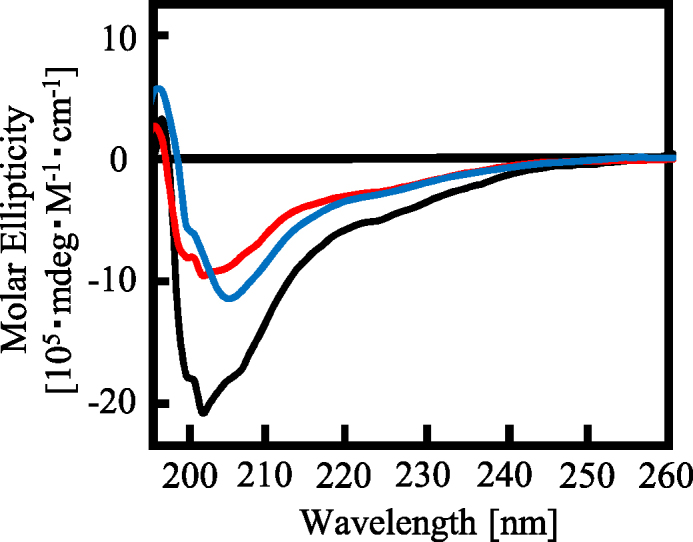
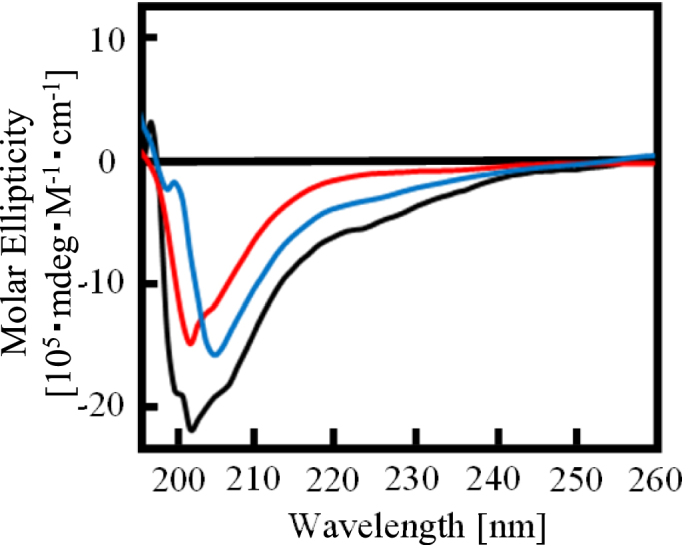
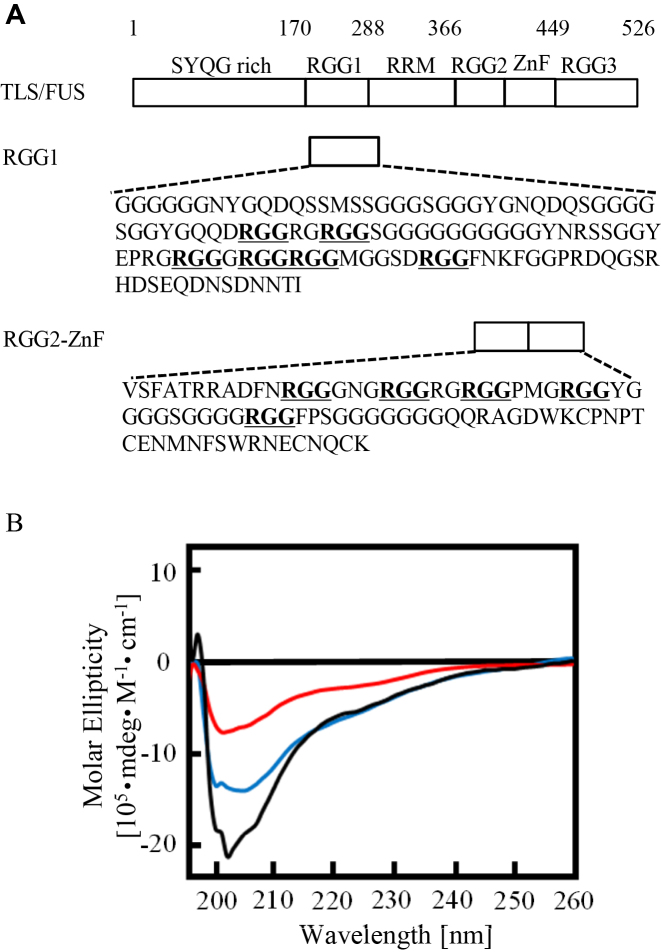
The RGG domain, defined as closely spaced Arg-Gly-Gly repeats, is a DNA and RNA-binding domain in various nucleic acid-binding proteins. Translocated in liposarcoma (TLS), which is also called FUS, is a protein with three RGG domains, RGG1, RGG2 and RGG3. TLS/FUS binding to G-quadruplex telomere DNA and telomeric repeat-containing RNA depends especially on RGG3, comprising Arg-Gly-Gly repeats with proline- and arginine-rich regions. So far, however, only non-specific DNA and RNA binding of TLS/FUS purified with buffers containing urea and KCl have been reported. Here, we demonstrate that protein purification using a buffer with high concentrations of urea and KCl decreases the G-quadruplex binding abilities of TLS/FUS and RGG3, and disrupts the β-spiral structure of RGG3. Moreover, the Arg-Gly-Gly repeat region in RGG3 by itself cannot form a stable β-spiral structure that binds to the G-quadruplex, because the proline- and arginine-rich regions induce the β-spiral structure and the G-quadruplex-binding ability of RGG3. Our findings suggest that the G-quadruplex-specific binding abilities of TLS/FUS require RGG3 with a β-spiral structure stabilized by adjacent proline- and arginine-regions.
RGG 结构域定义为紧密间隔的精氨酸-甘氨酸-甘氨酸重复序列,是各种核酸结合蛋白中的 DNA 和 RNA 结合结构域。易位性脂肪肉瘤(TLS),也称为 FUS,是一种具有三个 RGG 结构域的蛋白质,即 RGG1、RGG2 和 RGG3。TLS/FUS 与 G-四链体端粒 DNA 和富含端粒重复的 RNA 的结合特别依赖于 RGG3,它由富含脯氨酸和精氨酸的区域的精氨酸-甘氨酸-甘氨酸重复序列组成。然而,到目前为止,仅报道了使用含有尿素和 KCl 的缓冲液纯化的 TLS/FUS 具有非特异性的 DNA 和 RNA 结合能力。在这里,我们证明使用高浓度尿素和 KCl 的缓冲液进行蛋白质纯化会降低 TLS/FUS 和 RGG3 的 G-四链体结合能力,并破坏 RGG3 的 β-螺旋结构。此外,RGG3 自身的精氨酸-甘氨酸-甘氨酸重复区域不能形成稳定的结合 G-四链体的 β-螺旋结构,因为富含脯氨酸和精氨酸的区域诱导 RGG3 的 β-螺旋结构和 G-四链体结合能力。我们的研究结果表明,TLS/FUS 的 G-四链体特异性结合能力需要具有由相邻脯氨酸和精氨酸区域稳定的 β-螺旋结构的 RGG3。