Department of Physics, University of Illinois, Chicago, IL.
Department of Pathology, The University of Chicago, Chicago, IL.
J Gen Physiol. 2018 Jul 2;150(7):949-968. doi: 10.1085/jgp.201711868. Epub 2018 Jun 18.
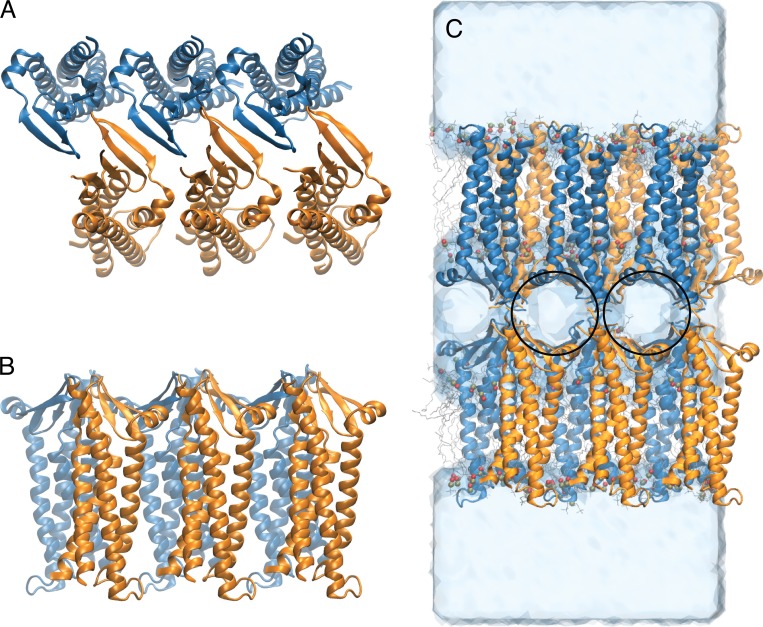
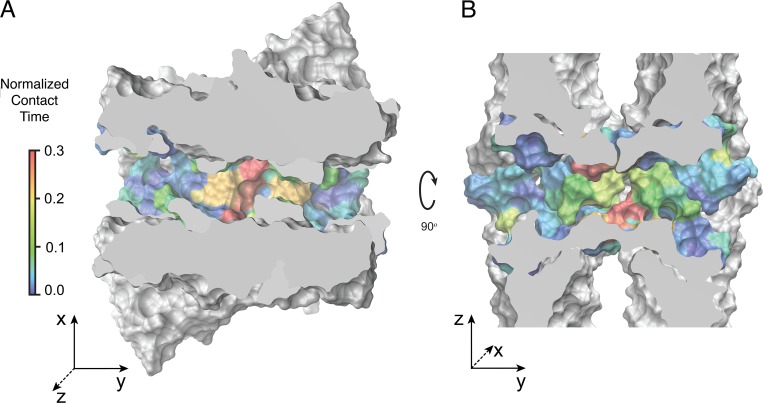
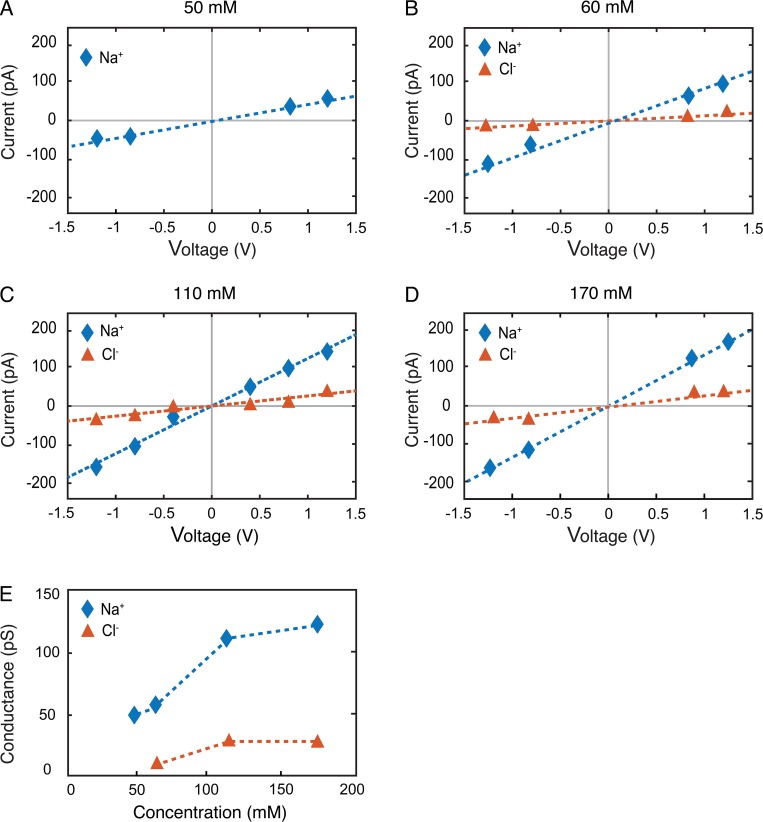
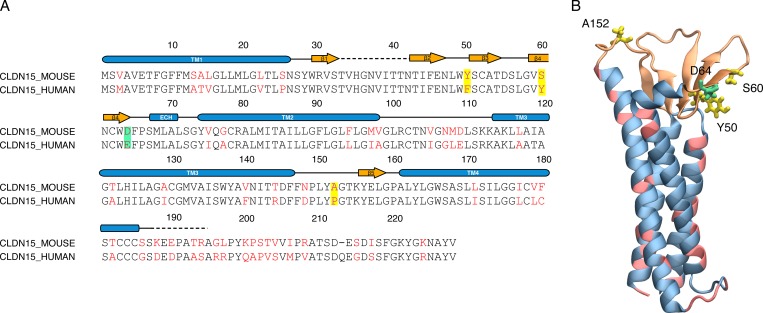
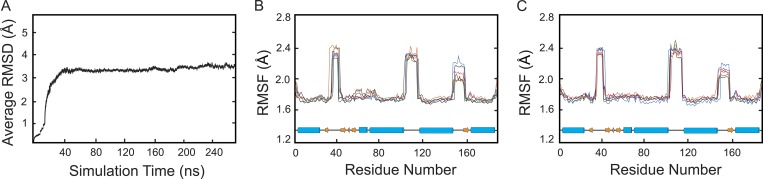
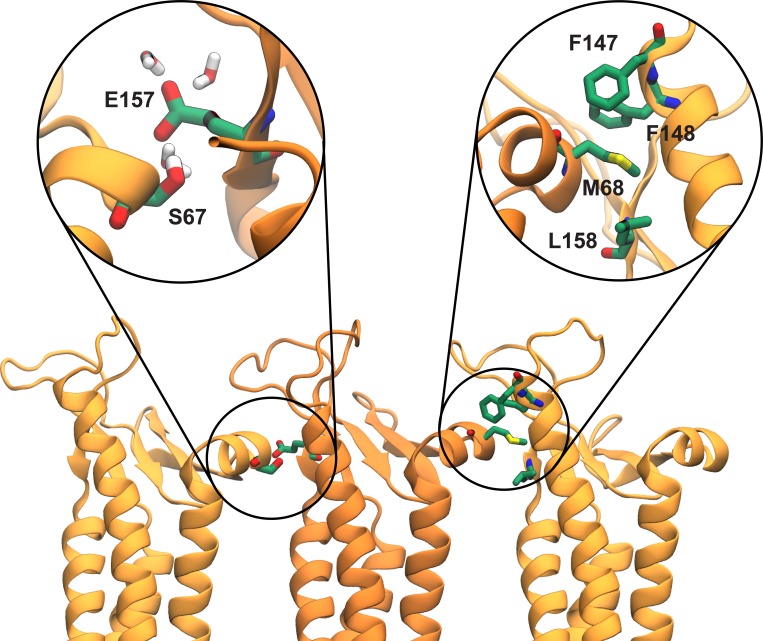
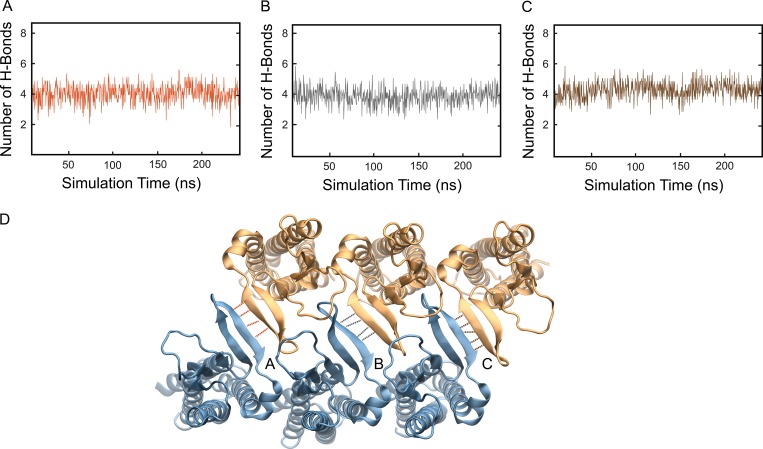
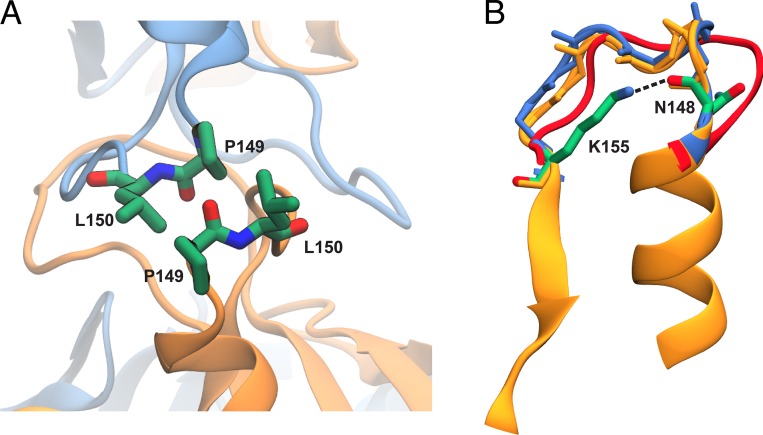
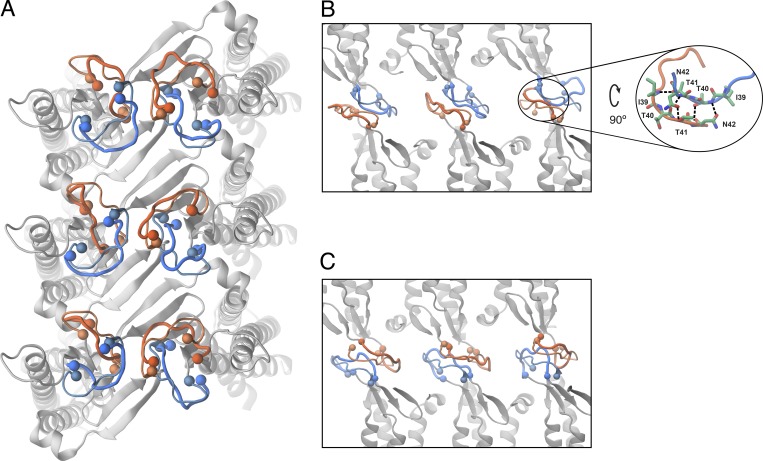
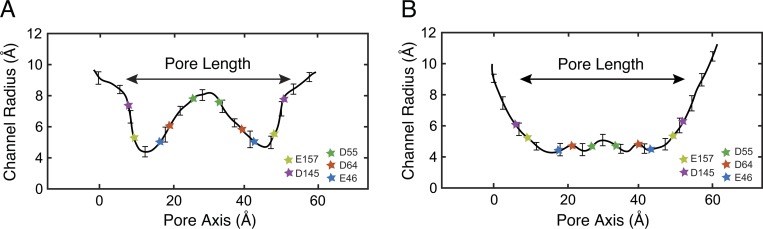
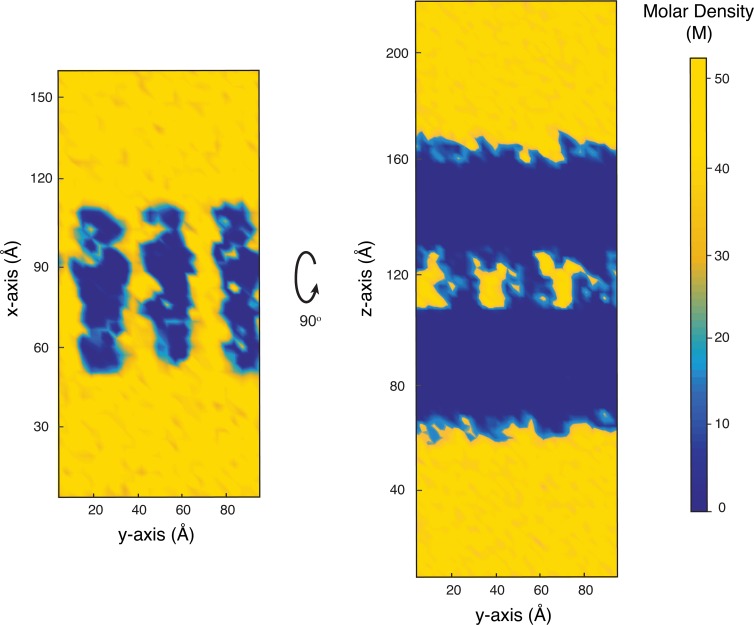
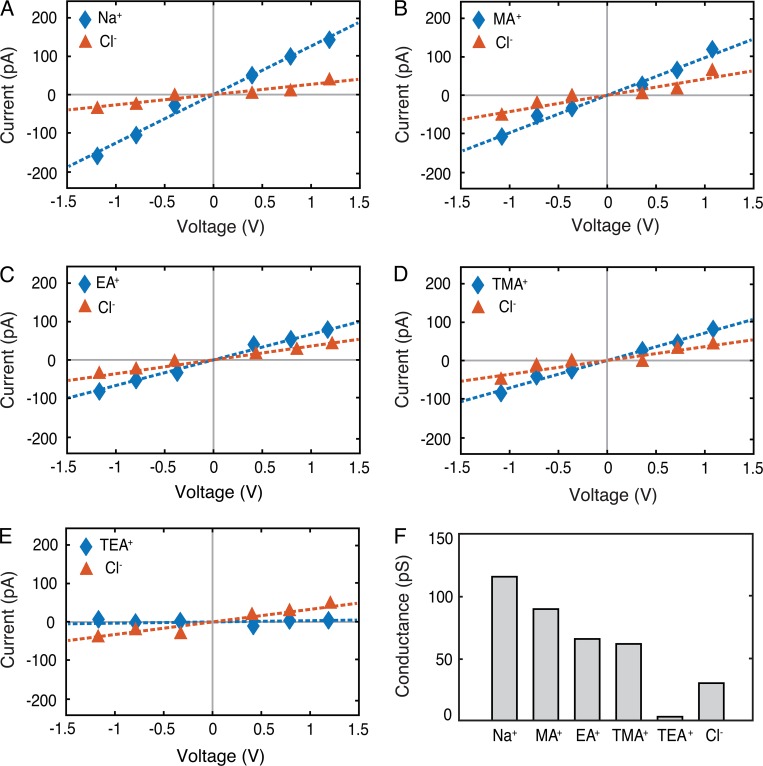
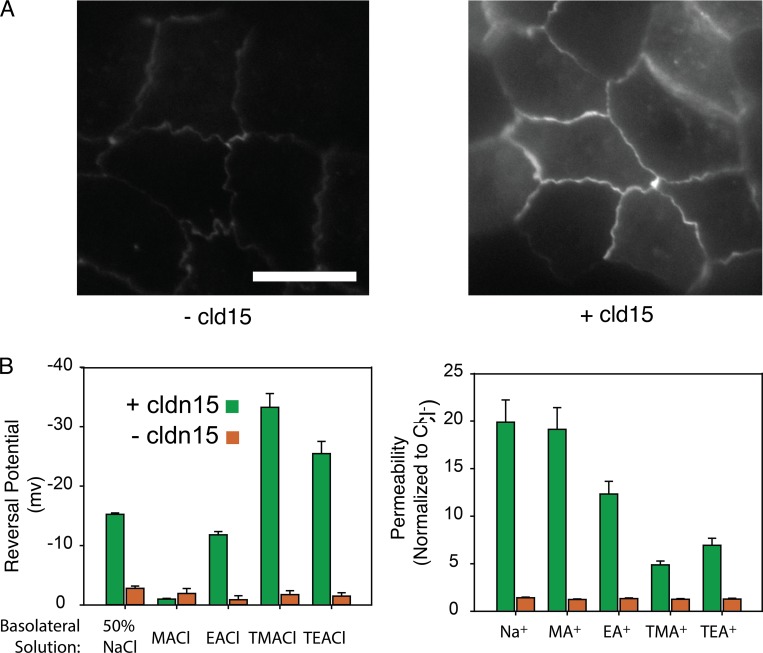
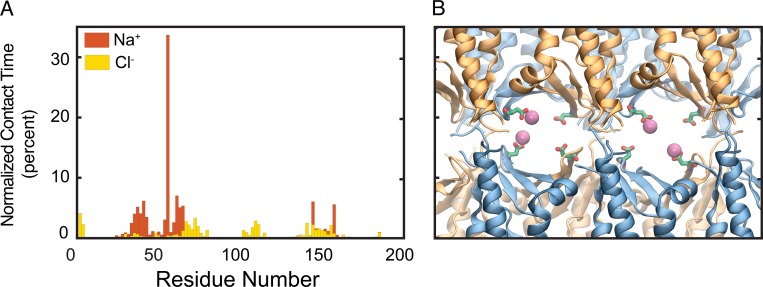
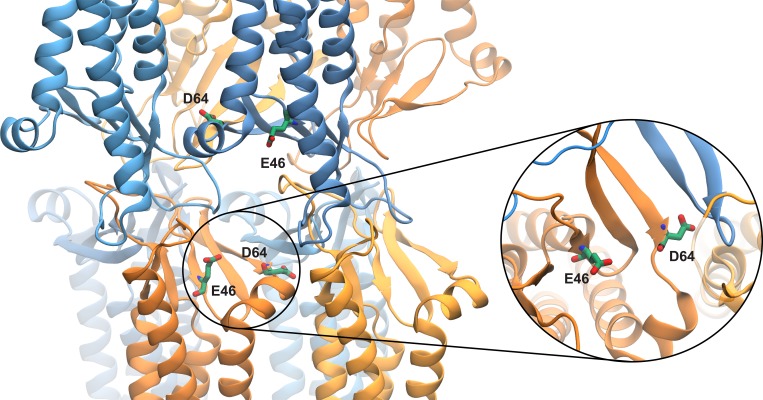
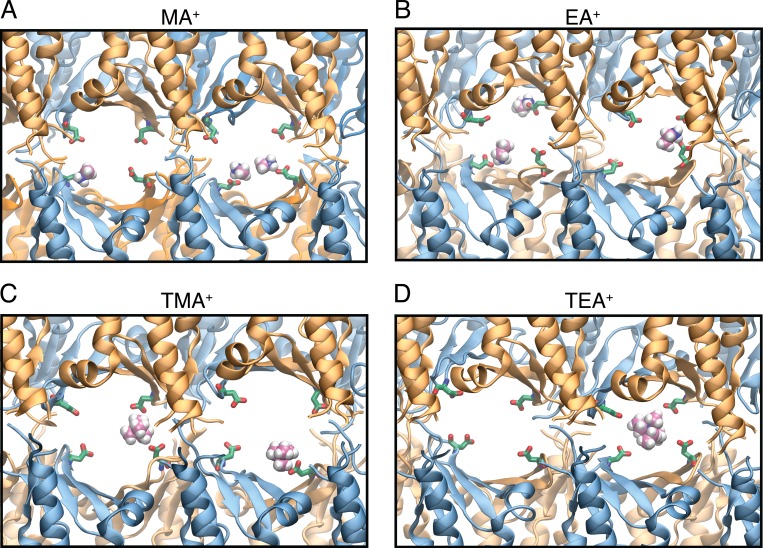
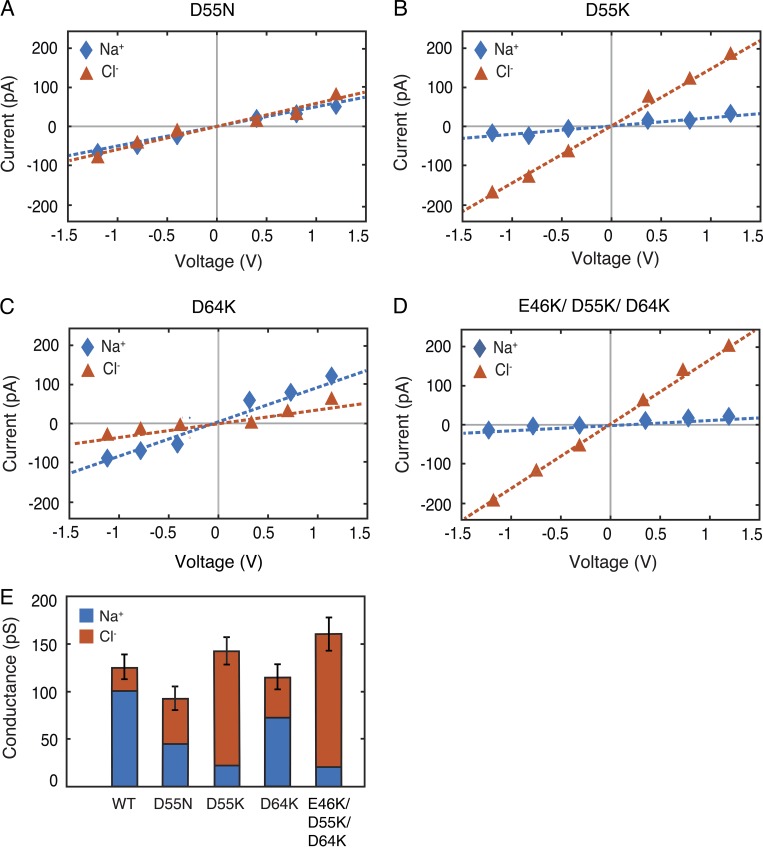
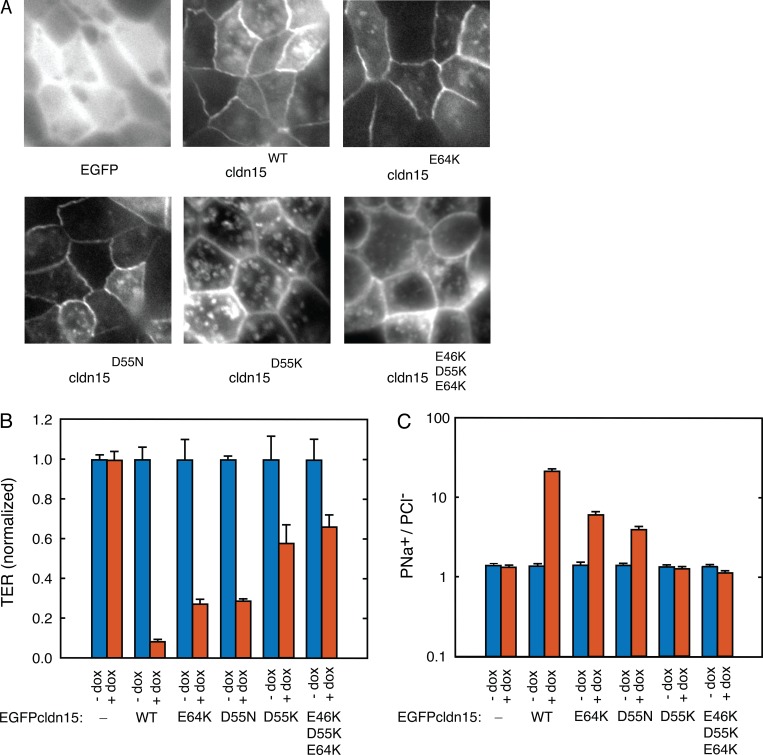
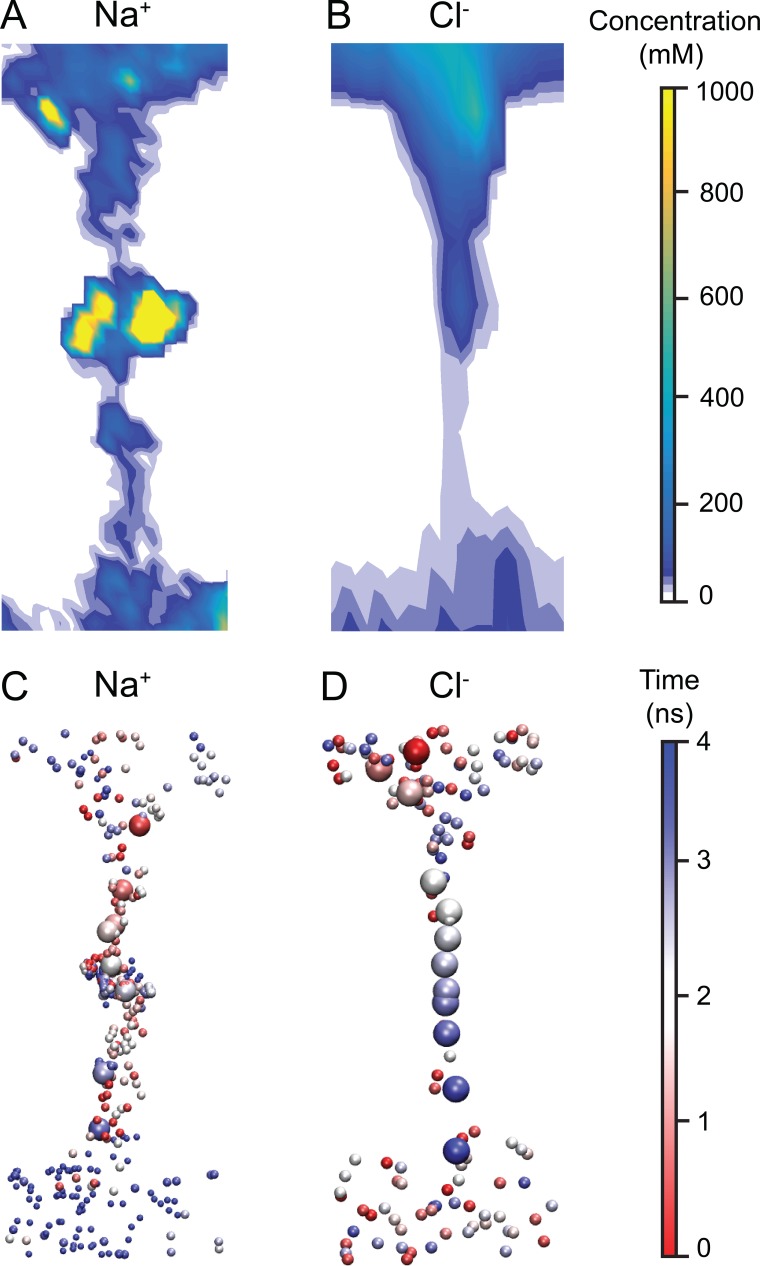
Tight junctions are macromolecular structures that traverse the space between adjacent cells in epithelia and endothelia. Members of the claudin family are known to determine tight junction permeability in a charge- and size-selective manner. Here, we use molecular dynamics simulations to build and refine an atomic model of claudin-15 channels and study its transport properties. Our simulations indicate that claudin-15 forms well-defined channels for ions and molecules and otherwise "seals" the paracellular space through hydrophobic interactions. Ionic currents, calculated from simulation trajectories of wild-type as well as mutant channels, reflect in vitro measurements. The simulations suggest that the selectivity filter is formed by a cage of four aspartic acid residues (D55), contributed by four claudin-15 molecules, which creates a negative electrostatic potential to favor cation flux over anion flux. Charge reversal or charge ablation mutations of D55 significantly reduce cation permeability in silico and in vitro, whereas mutations of other negatively charged pore amino acid residues have a significantly smaller impact on channel permeability and selectivity. The simulations also indicate that water and small ions can pass through the channel, but larger cations, such as tetramethylammonium, do not traverse the pore. Thus, our model provides an atomic view of claudin channels, their transport function, and a potential three-dimensional organization of its selectivity filter.
紧密连接是横跨上皮细胞和内皮细胞之间的空间的大分子结构。已知 claudin 家族的成员以电荷和大小选择性的方式决定紧密连接的通透性。在这里,我们使用分子动力学模拟来构建和改进 claudin-15 通道的原子模型,并研究其传输特性。我们的模拟表明,claudin-15 形成了离子和分子的明确通道,并且通过疏水性相互作用“密封”了细胞旁空间。从野生型和突变通道的模拟轨迹计算出的离子电流反映了体外测量结果。模拟表明,选择性过滤器由四个天冬氨酸残基(D55)组成的笼形成,由四个 claudin-15 分子贡献,形成负静电势以有利于阳离子通量而不是阴离子通量。D55 的电荷反转或电荷消融突变在计算机模拟和体外显著降低了阳离子通透性,而其他带负电荷的孔氨基酸残基的突变对通道通透性和选择性的影响要小得多。模拟还表明,水和小离子可以通过通道,但像四甲基铵这样的较大阳离子不能穿过孔。因此,我们的模型提供了 claudin 通道的原子视图、它们的传输功能以及其选择性过滤器的潜在三维组织。