Division of Infectious Diseases, Department of Internal Medicine, University of Michigan, Ann Arbor, Michigan, United States of America.
Department of Microbiology and Immunology, University of Michigan, Ann Arbor, Michigan, United States of America.
PLoS Biol. 2018 Jun 28;16(6):e2006459. doi: 10.1371/journal.pbio.2006459. eCollection 2018 Jun.
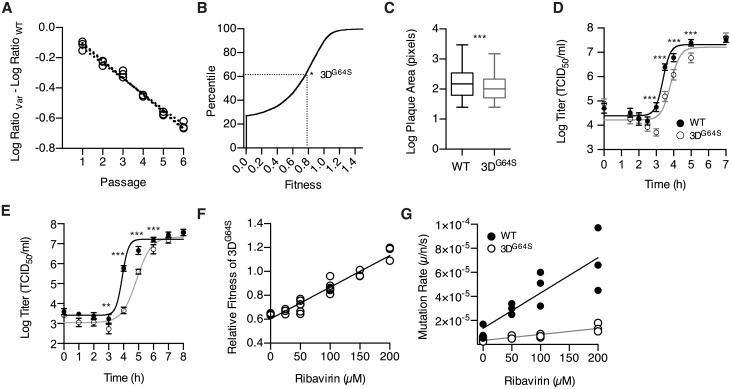
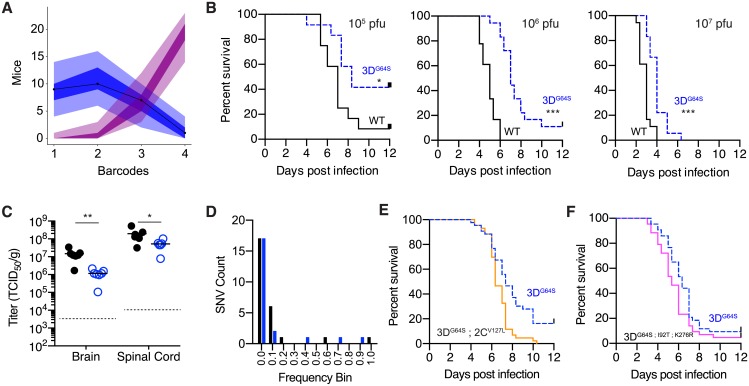
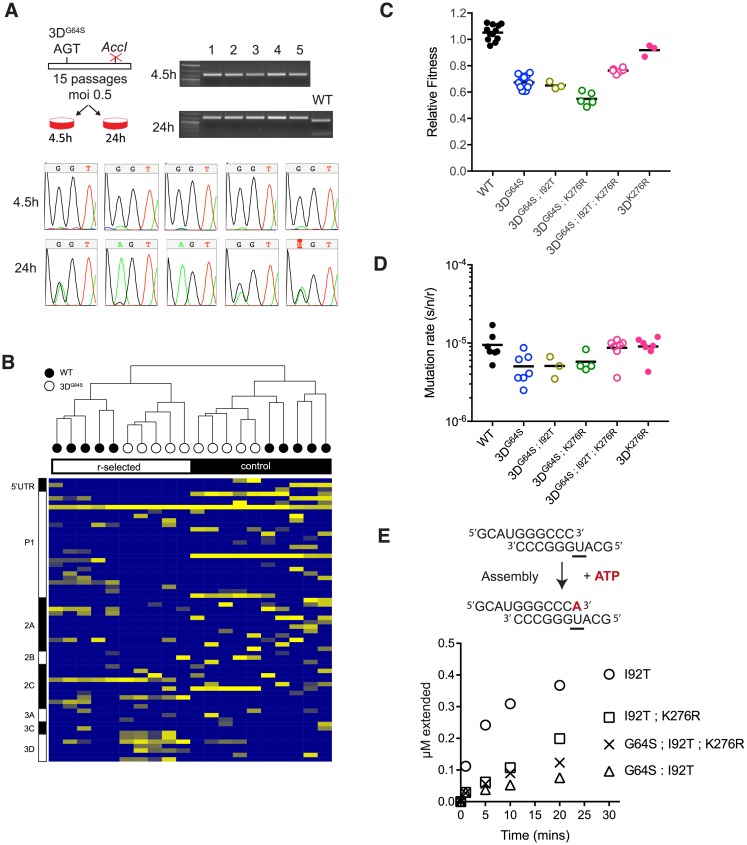
Mutation rates can evolve through genetic drift, indirect selection due to genetic hitchhiking, or direct selection on the physicochemical cost of high fidelity. However, for many systems, it has been difficult to disentangle the relative impact of these forces empirically. In RNA viruses, an observed correlation between mutation rate and virulence has led many to argue that their extremely high mutation rates are advantageous because they may allow for increased adaptability. This argument has profound implications because it suggests that pathogenesis in many viral infections depends on rare or de novo mutations. Here, we present data for an alternative model whereby RNA viruses evolve high mutation rates as a byproduct of selection for increased replicative speed. We find that a poliovirus antimutator, 3DG64S, has a significant replication defect and that wild-type (WT) and 3DG64S populations have similar adaptability in 2 distinct cellular environments. Experimental evolution of 3DG64S under selection for replicative speed led to reversion and compensation of the fidelity phenotype. Mice infected with 3DG64S exhibited delayed morbidity at doses well above the lethal level, consistent with attenuation by slower growth as opposed to reduced mutational supply. Furthermore, compensation of the 3DG64S growth defect restored virulence, while compensation of the fidelity phenotype did not. Our data are consistent with the kinetic proofreading model for biosynthetic reactions and suggest that speed is more important than accuracy. In contrast with what has been suggested for many RNA viruses, we find that within-host spread is associated with viral replicative speed and not standing genetic diversity.
突变率可以通过遗传漂变、由于遗传连锁而产生的间接选择,或对高保真度的物理化学成本的直接选择而进化。然而,对于许多系统来说,很难从经验上区分这些力量的相对影响。在 RNA 病毒中,突变率与毒力之间的观察到的相关性导致许多人认为它们极高的突变率是有利的,因为它们可能允许更高的适应性。这个论点具有深远的意义,因为它表明许多病毒感染的发病机制取决于罕见的或新出现的突变。在这里,我们提出了一个替代模型的数据,即 RNA 病毒通过选择增加复制速度而进化出高突变率。我们发现,脊髓灰质炎病毒的抗突变体 3DG64S 具有明显的复制缺陷,并且野生型 (WT) 和 3DG64S 群体在 2 种不同的细胞环境中具有相似的适应性。在选择复制速度的条件下对 3DG64S 进行实验进化导致保真度表型的回复和补偿。在远远高于致死剂量的剂量下感染 3DG64S 的小鼠表现出发病延迟,这与生长速度较慢而不是突变供应减少导致的衰减一致。此外,3DG64S 生长缺陷的补偿恢复了毒力,而保真度表型的补偿则没有。我们的数据与生物合成反应的动力学校对模型一致,并表明速度比准确性更重要。与许多 RNA 病毒所暗示的相反,我们发现,在宿主内传播与病毒复制速度有关,而不是与遗传多样性有关。