MRC Laboratory of Molecular Biology, Cambridge, United Kingdom.
Instituto de Biomedicina de Valencia (IBV-CSIC), Valencia, Spain.
Elife. 2018 Nov 30;7:e39273. doi: 10.7554/eLife.39273.
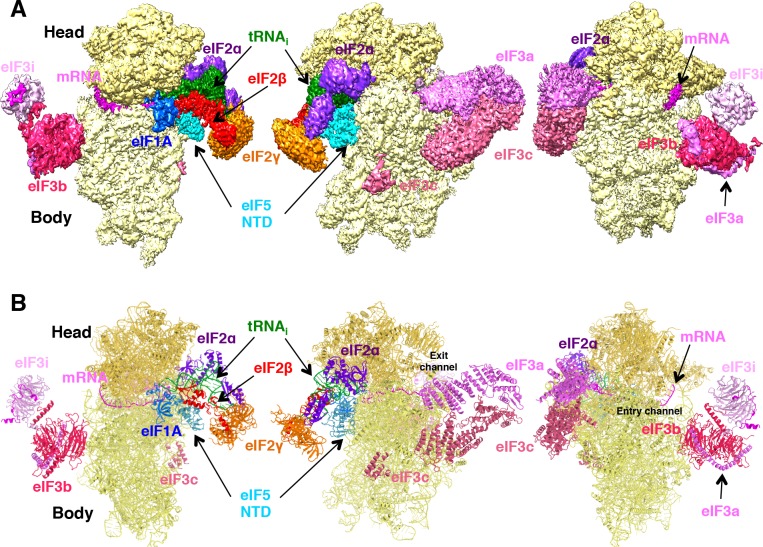
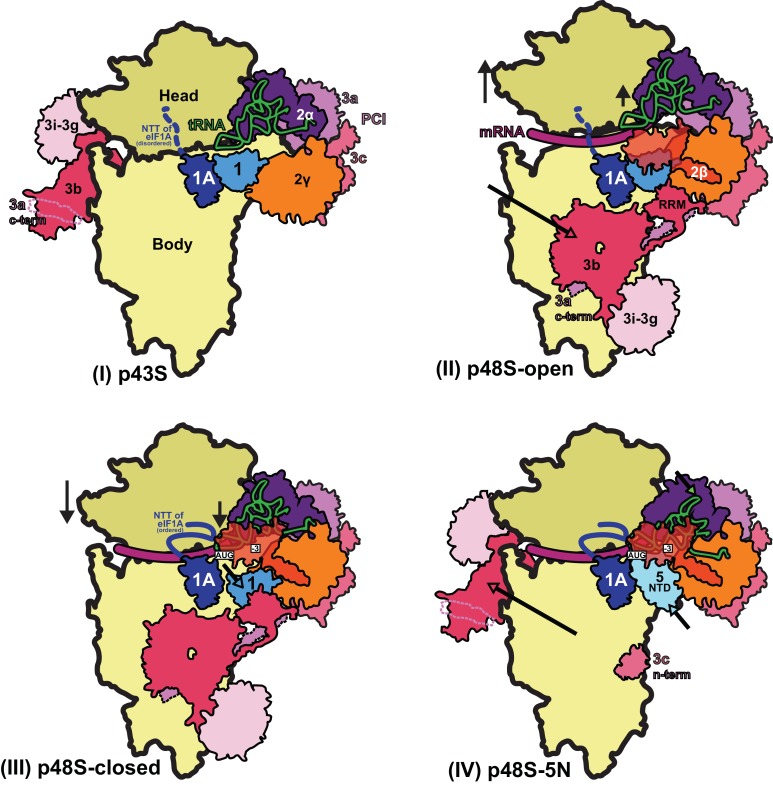
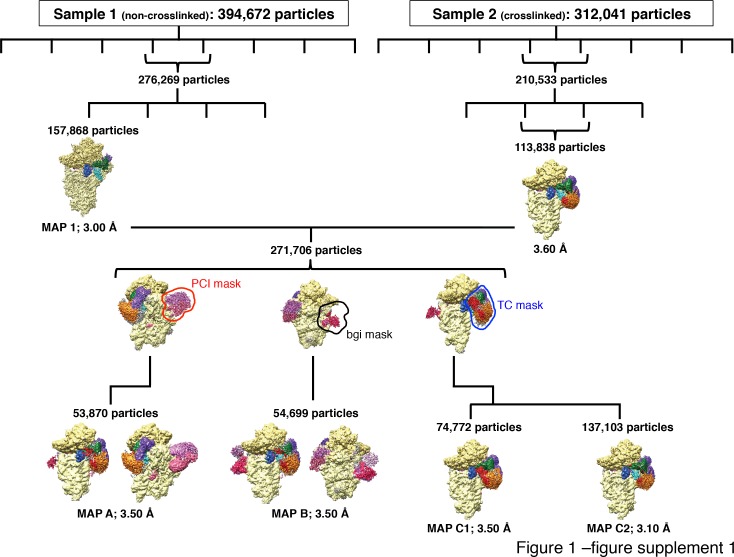
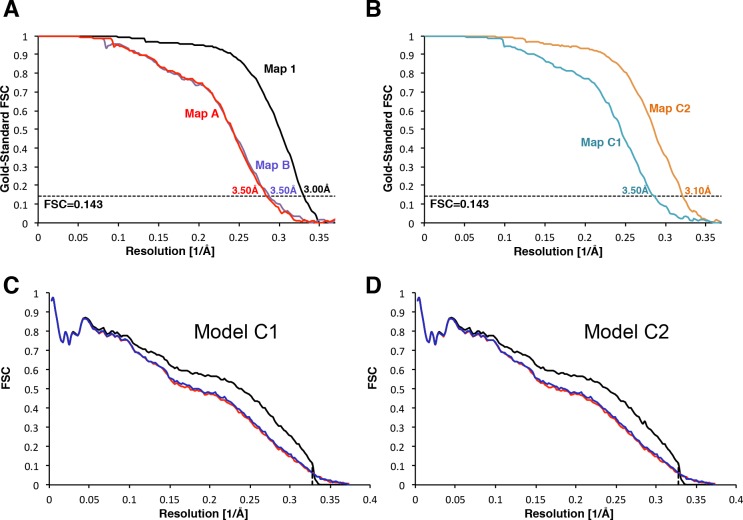
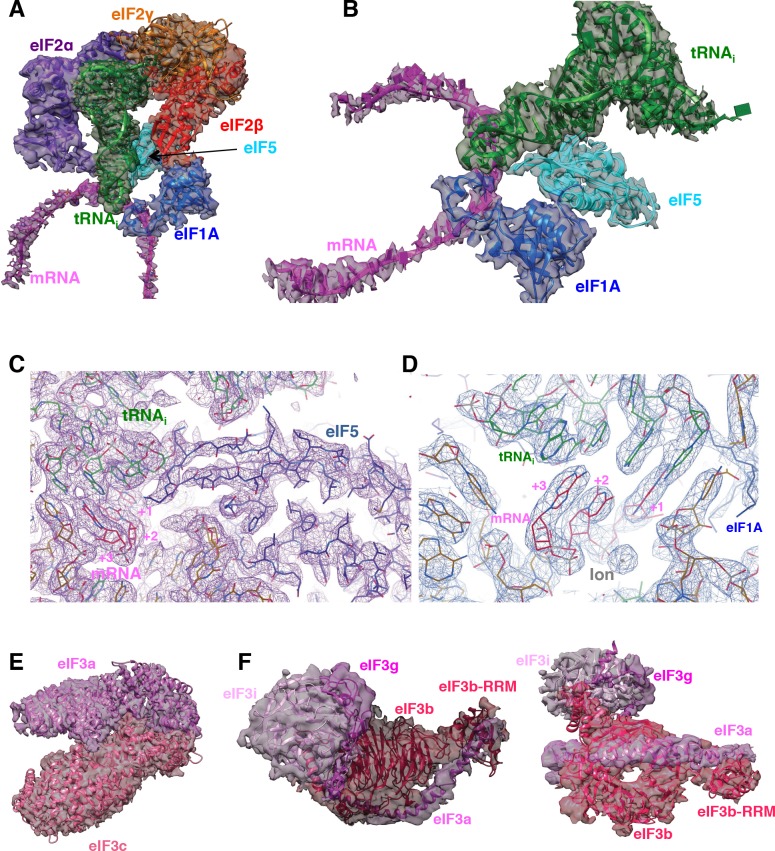
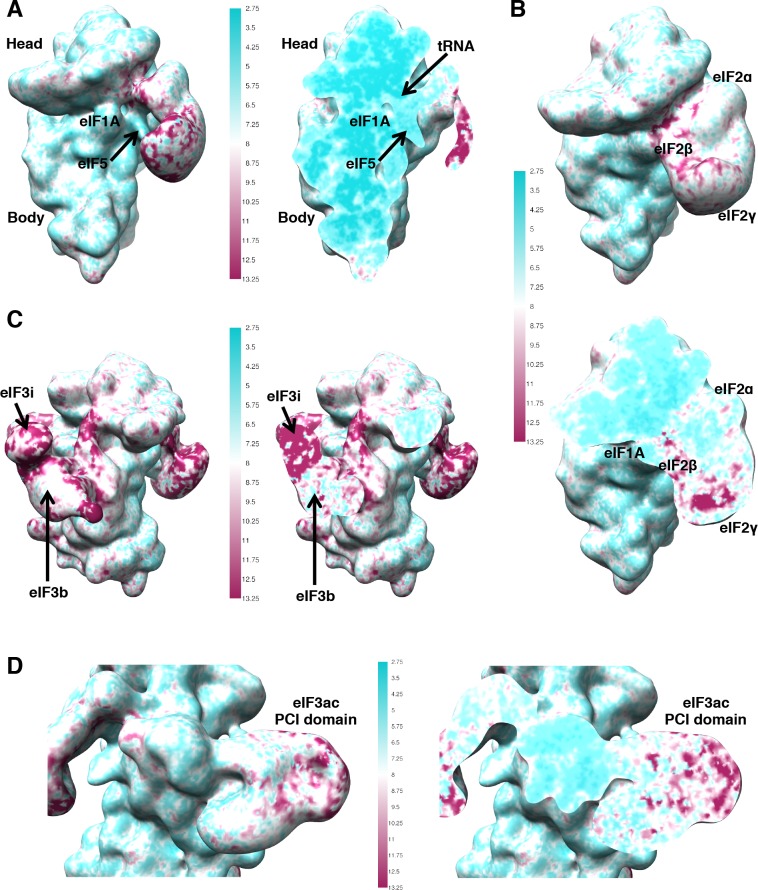
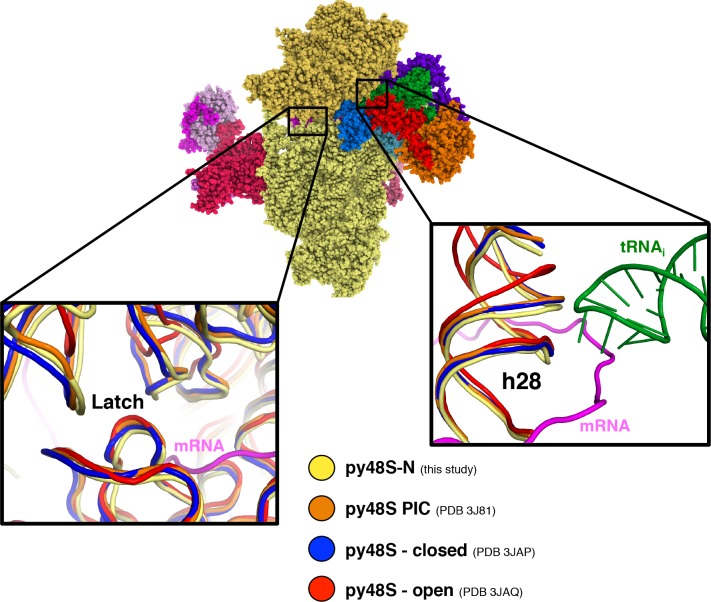
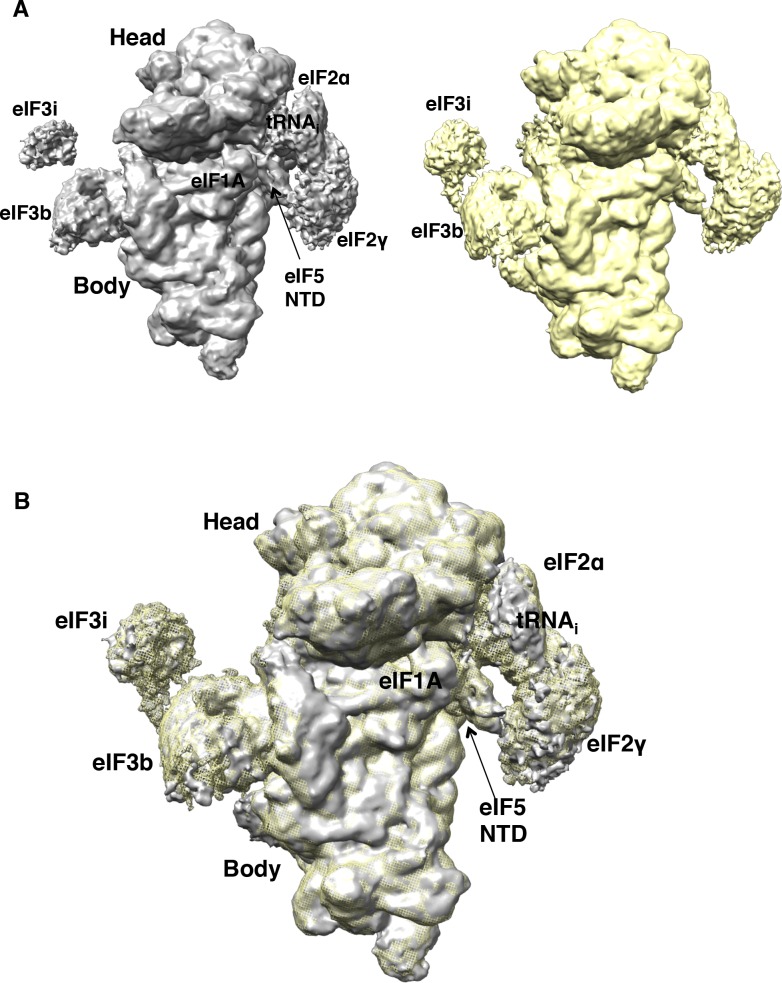
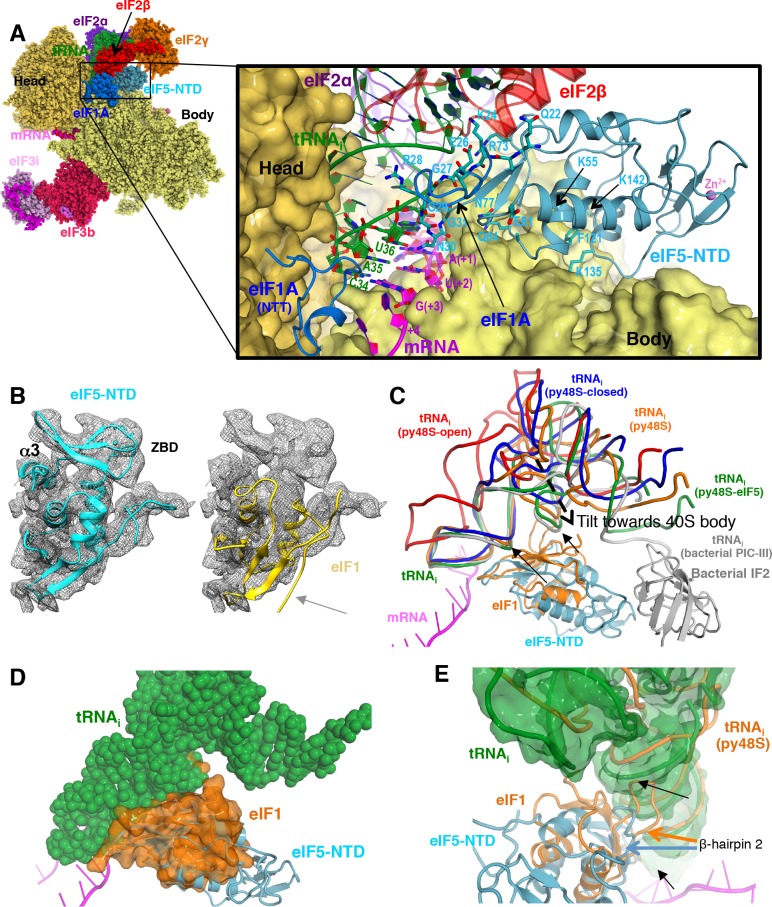
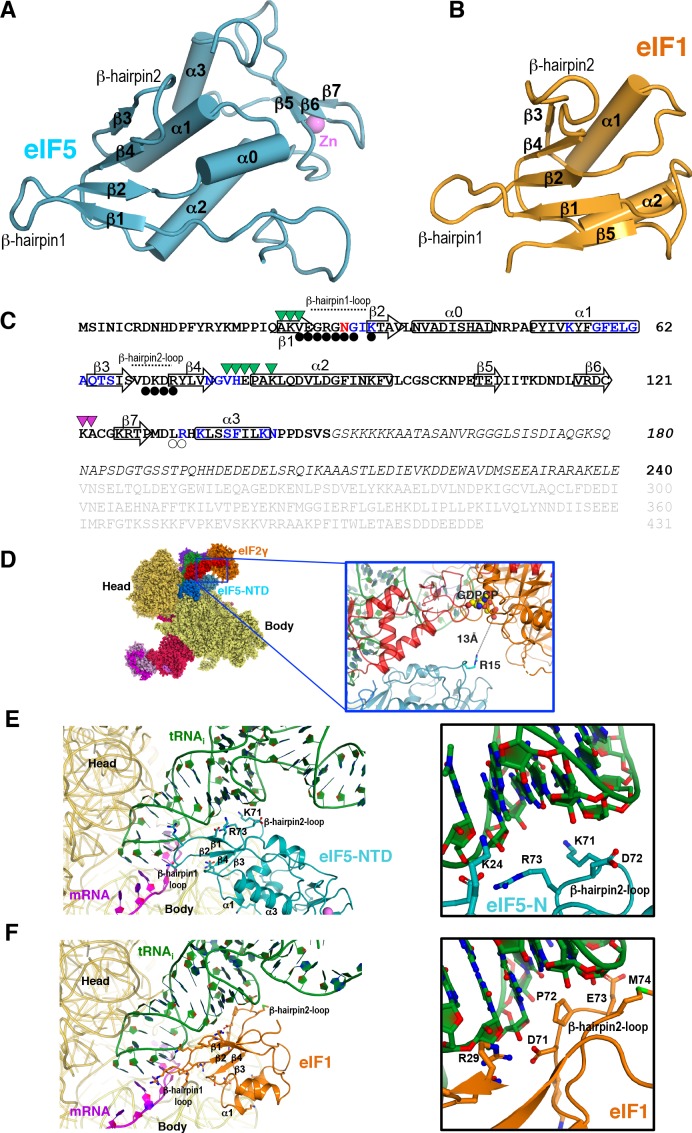
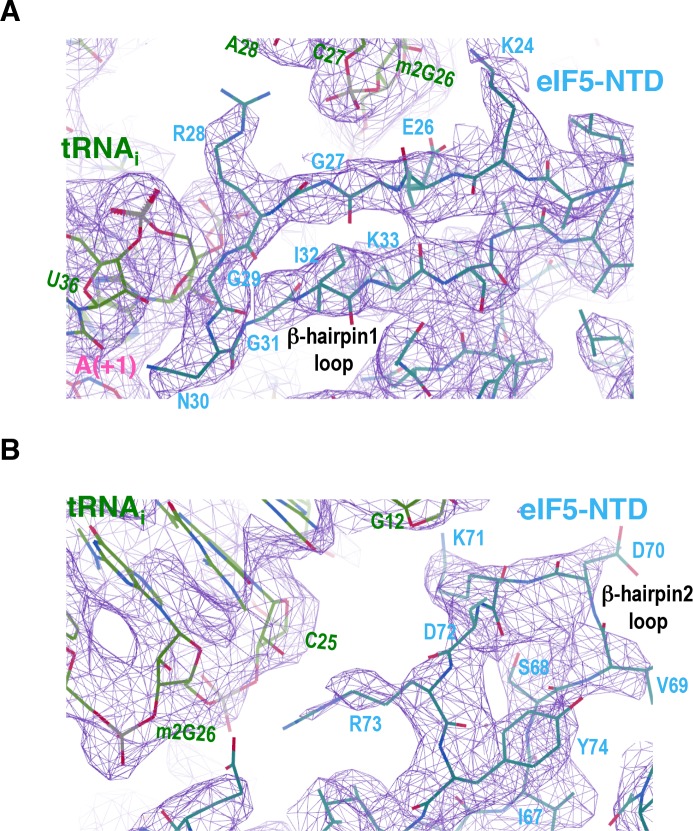
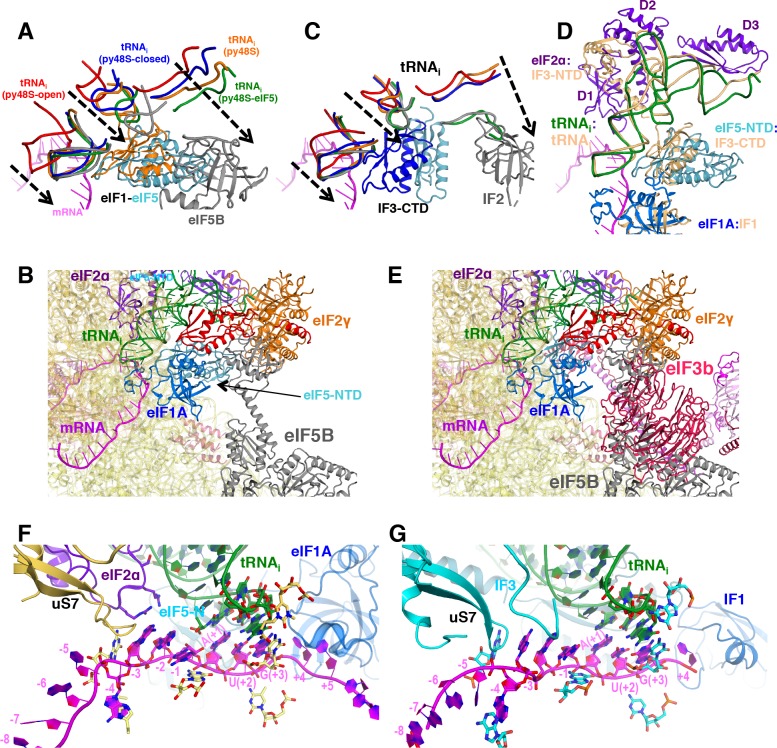
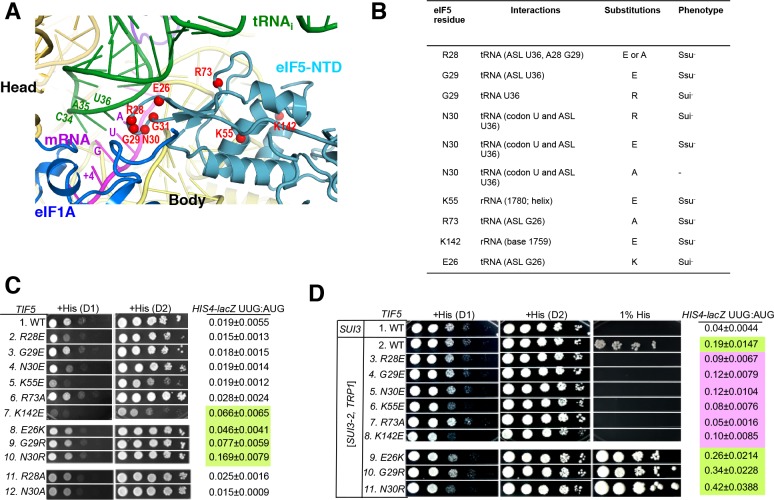
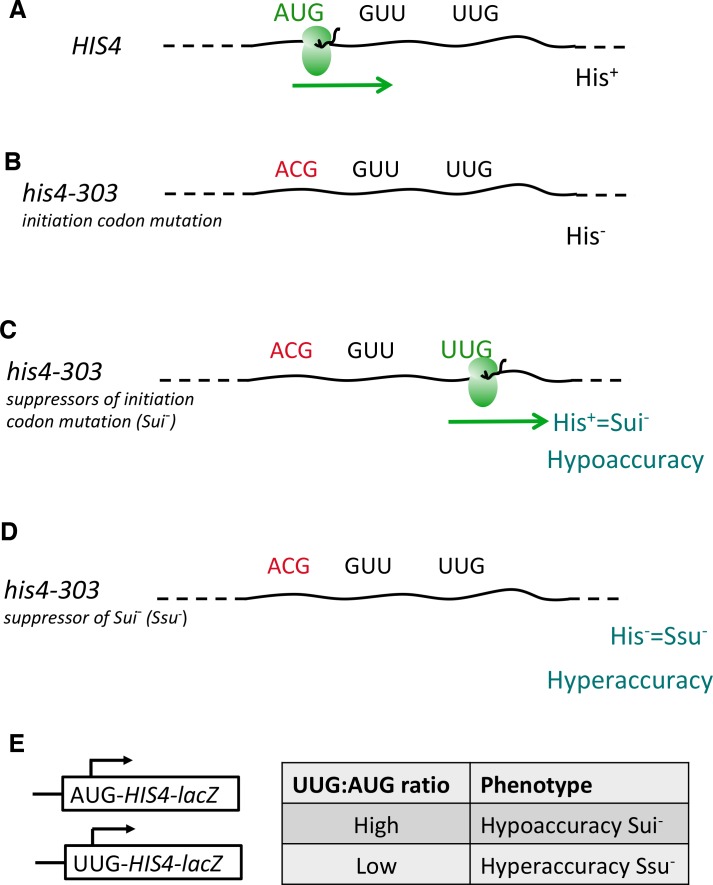
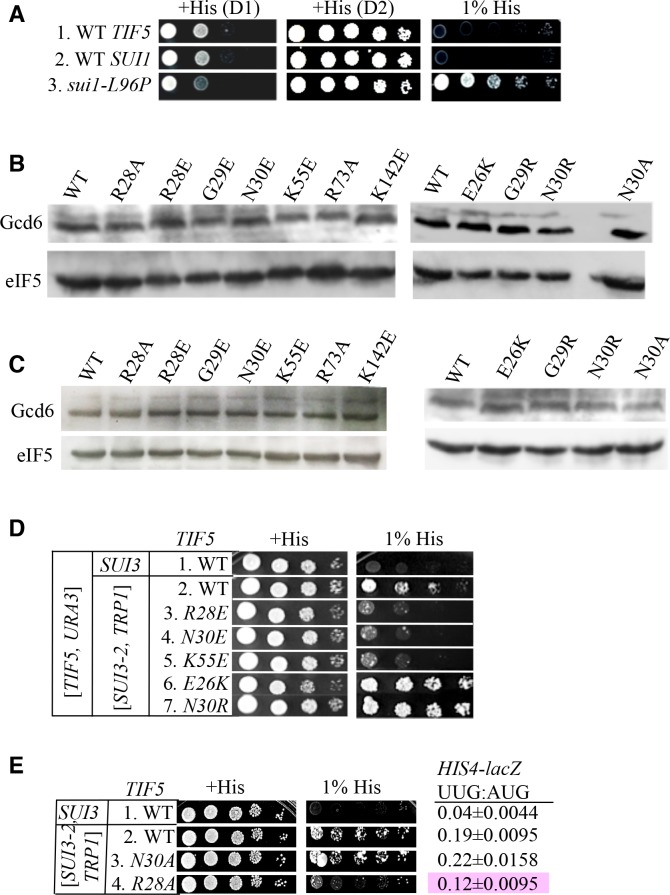
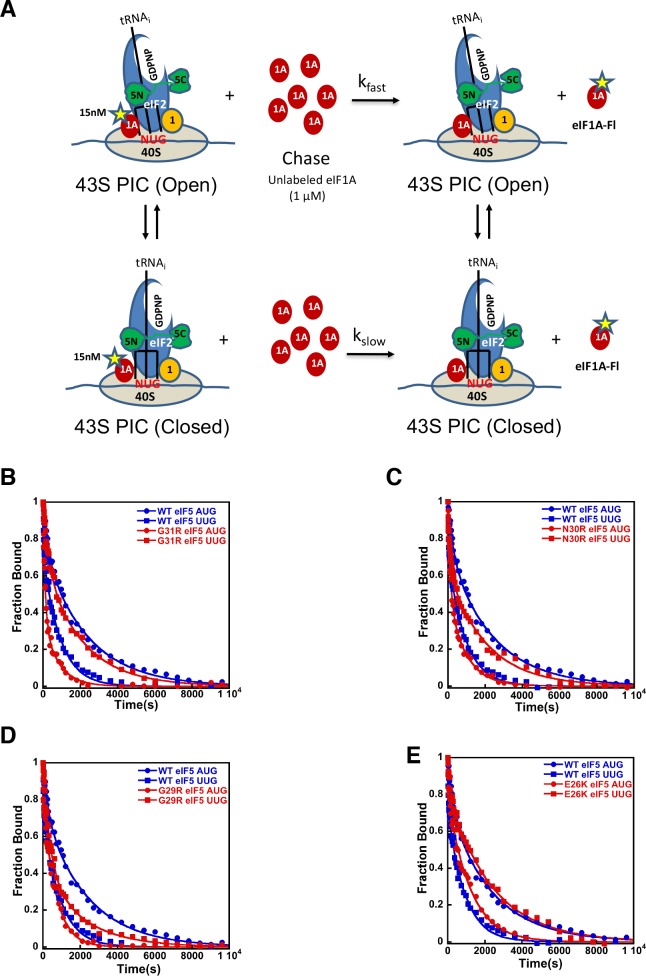
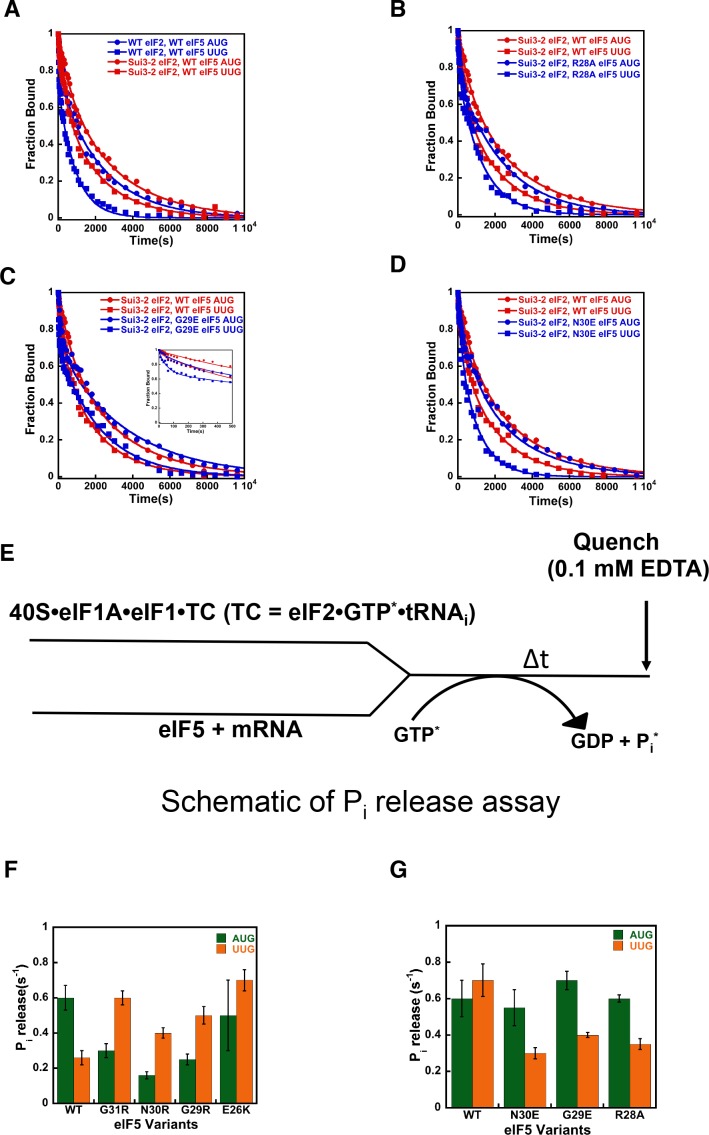
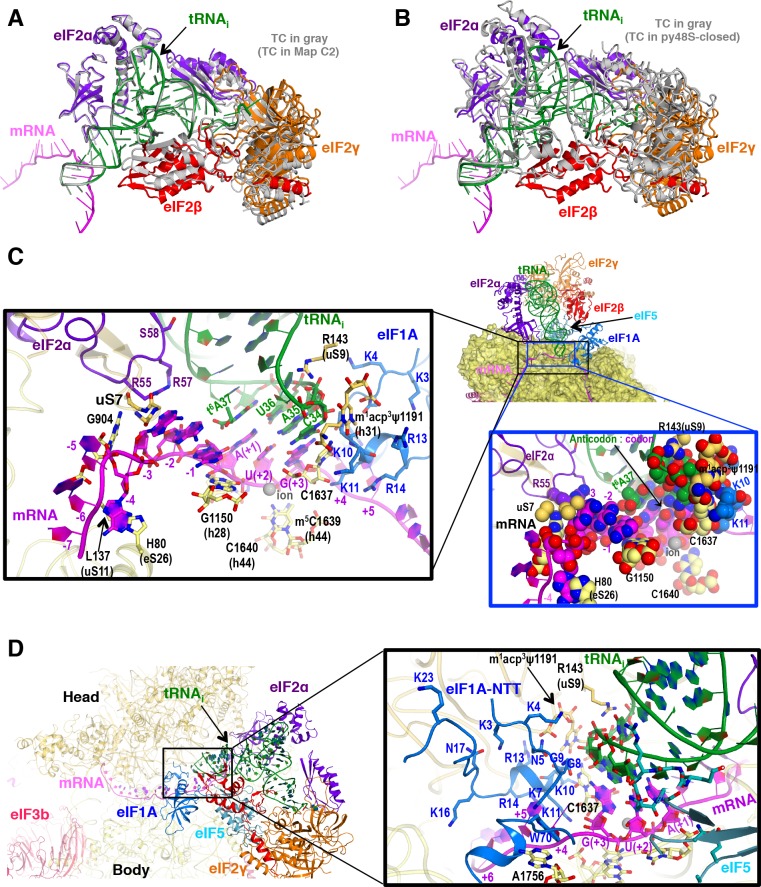
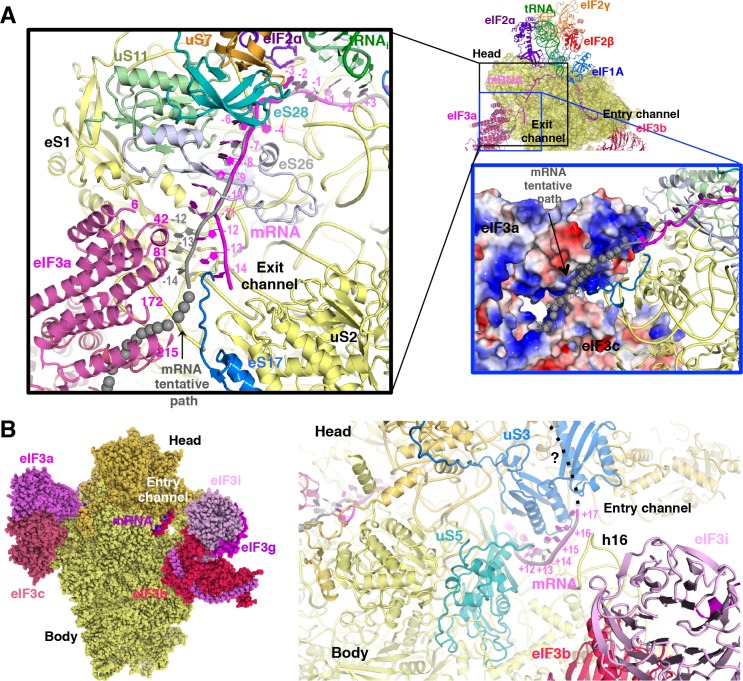
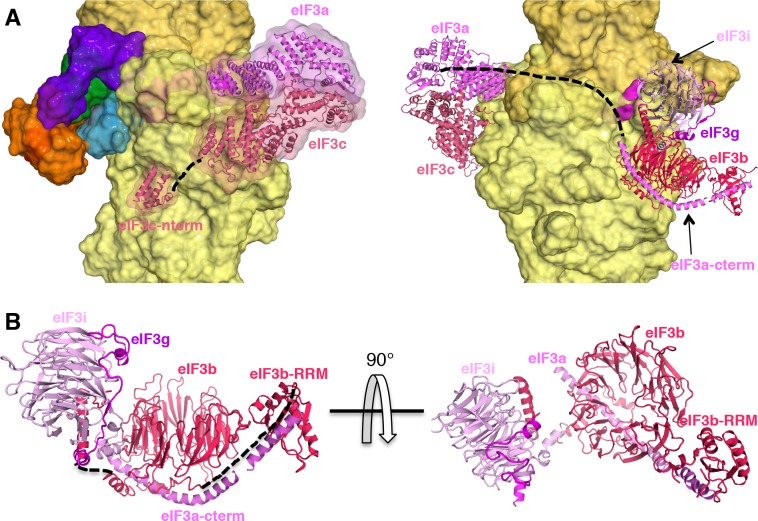
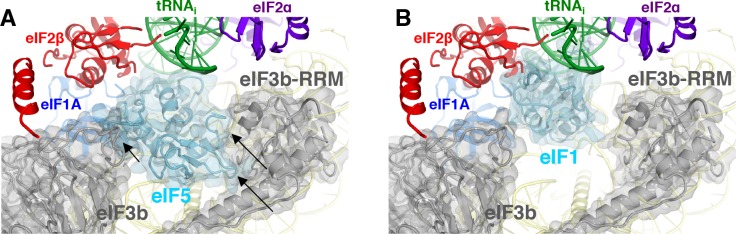
In eukaryotic translation initiation, AUG recognition of the mRNA requires accommodation of Met-tRNA in a 'P' state, which is antagonized by the factor eIF1. eIF5 is a GTPase activating protein (GAP) of eIF2 that additionally promotes stringent AUG selection, but the molecular basis of its dual function was unknown. We present a cryo-electron microscopy (cryo-EM) reconstruction of a yeast 48S pre-initiation complex (PIC), at an overall resolution of 3.0 Å, featuring the N-terminal domain (NTD) of eIF5 bound to the 40S subunit at the location vacated by eIF1. eIF5 interacts with and allows a more accommodated orientation of Met-tRNA. Substitutions of eIF5 residues involved in the eIF5-NTD/tRNA interaction influenced initiation at near-cognate UUG codons and the closed/open PIC conformation in vitro, consistent with direct stabilization of the codon:anticodon duplex by the wild-type eIF5-NTD. The present structure reveals the basis for a key role of eIF5 in start-codon selection.
在真核翻译起始中,AUG 对 mRNA 的识别需要使 Met-tRNA 适应“P”状态,而这一状态会受到因子 eIF1 的拮抗。eIF5 是 eIF2 的 GTP 酶激活蛋白 (GAP),它还促进严格的 AUG 选择,但它的双重功能的分子基础尚不清楚。我们展示了一个酵母 48S 起始前复合物 (PIC) 的冷冻电镜 (cryo-EM) 重建,整体分辨率为 3.0 Å,其中 eIF5 的 N 端结构域 (NTD) 结合在 40S 亚基上,占据了 eIF1 离开的位置。eIF5 与 Met-tRNA 相互作用,并允许其更适应的取向。与 eIF5-NTD/tRNA 相互作用相关的 eIF5 残基的取代会影响近同功 UUG 密码子的起始和体外封闭/开放 PIC 构象,这与野生型 eIF5-NTD 直接稳定密码子:反密码子二聚体一致。目前的结构揭示了 eIF5 在起始密码子选择中关键作用的基础。