Center for Global Infectious Disease Research, Seattle Children's Research Institute, Seattle, WA, United States.
Institute for Systems Biology, Seattle, WA, United States.
Front Cell Infect Microbiol. 2018 Nov 27;8:413. doi: 10.3389/fcimb.2018.00413. eCollection 2018.
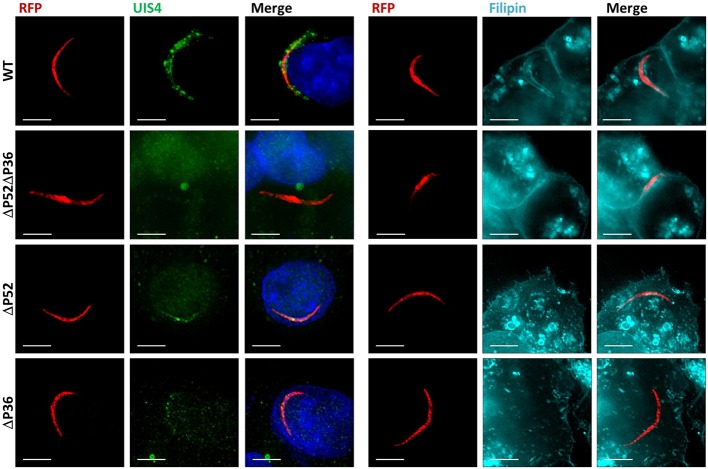
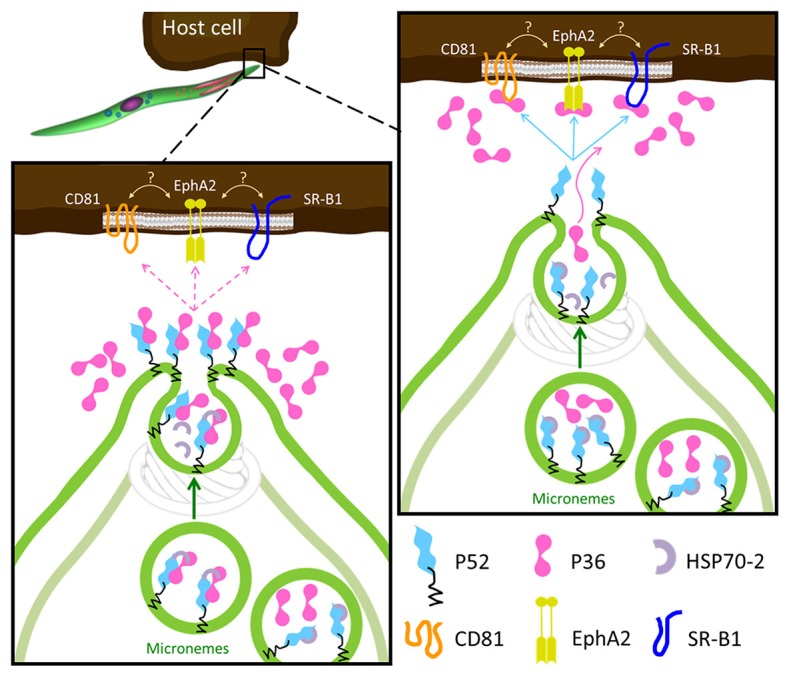
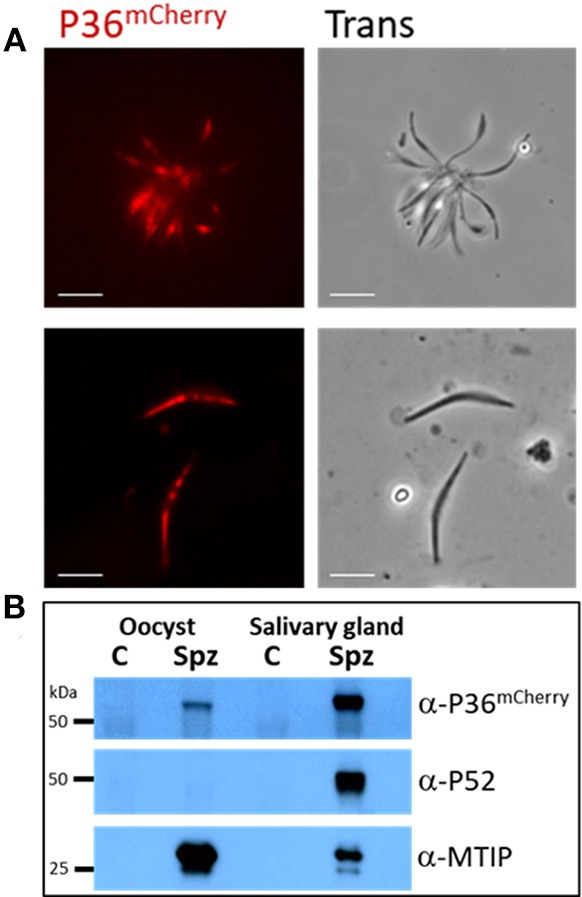
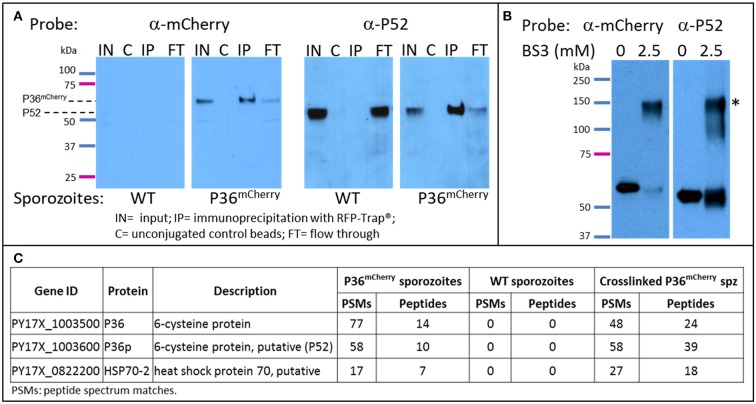
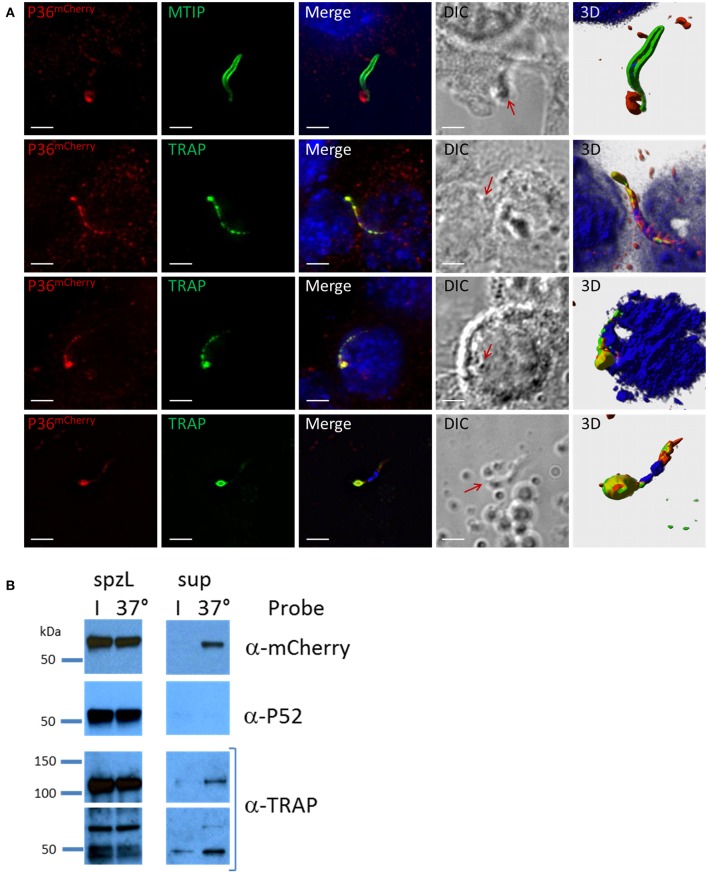
Within the liver, sporozoites traverse cells searching for a "suitable" hepatocyte, invading these cells through a process that results in the formation of a parasitophorous vacuole (PV), within which the parasite undergoes intracellular replication as a liver stage. It was previously established that two members of the s48/45 protein family, P36 and P52, are essential for productive invasion of host hepatocytes by sporozoites as their simultaneous deletion results in growth-arrested parasites that lack a PV. Recent studies point toward a pathway of entry possibly involving the interaction of P36 with hepatocyte receptors EphA2, CD81, and SR-B1. However, the relationship between P36 and P52 during sporozoite invasion remains unknown. Here we show that parasites with a single or gene deletion each lack a PV after hepatocyte invasion, thereby pheno-copying the lack of a PV observed for the / dual gene deletion parasite line. This indicates that both proteins are equally important in the establishment of a PV and act in the same pathway. We created a P36 tagged parasite line that allowed us to visualize the subcellular localization of P36 and found that it partially co-localizes with P52 in the sporozoite secretory microneme organelles. Furthermore, through co-immunoprecipitation studies , we determined that P36 and P52 form a protein complex in sporozoites, indicating a concerted function for both proteins within the PV formation pathway. However, upon sporozoite stimulation, only P36 was released as a secreted protein while P52 was not. Our results support a model in which the putatively glycosylphosphatidylinositol (GPI)-anchored P52 may serve as a scaffold to facilitate the interaction of secreted P36 with the host cell during sporozoite invasion of hepatocytes.
在肝脏内,子孢子穿过细胞寻找“合适”的肝细胞,通过导致形成寄生泡(PV)的过程侵入这些细胞,在其中寄生虫作为肝期经历细胞内复制。先前已经确立,s48/45 蛋白家族的两个成员,P36 和 P52,对于子孢子对宿主肝细胞的有效侵入是必需的,因为它们的同时缺失导致生长停滞的寄生虫缺乏 PV。最近的研究表明,可能涉及 P36 与肝细胞受体 EphA2、CD81 和 SR-B1 相互作用的进入途径。然而,在子孢子侵入期间 P36 和 P52 之间的关系仍然未知。在这里,我们显示每个具有单个 或 基因缺失的寄生虫在侵入肝细胞后都缺乏 PV,从而模拟了观察到的 / 双基因缺失寄生虫系缺乏 PV 的现象。这表明这两种蛋白质在 PV 的建立中同样重要,并且作用于相同的途径。我们创建了一个 P36 标记的寄生虫系,使我们能够可视化 P36 的亚细胞定位,并发现它部分与 P52 在子孢子分泌微纳细胞器中共定位。此外,通过共免疫沉淀研究,我们确定 P36 和 P52 在子孢子中形成蛋白质复合物,表明这两种蛋白质在 PV 形成途径中具有协同功能。然而,在子孢子刺激后,只有 P36 作为分泌蛋白释放,而 P52 则没有。我们的结果支持了这样一种模型,即假定糖基磷脂酰肌醇(GPI)锚定的 P52 可能作为支架,促进分泌的 P36 在子孢子侵入肝细胞期间与宿主细胞的相互作用。