Bourguet Pierre, de Bossoreille Stève, López-González Leticia, Pouch-Pélissier Marie-Noëlle, Gómez-Zambrano Ángeles, Devert Anthony, Pélissier Thierry, Pogorelcnik Romain, Vaillant Isabelle, Mathieu Olivier
Génétique Reproduction et Développement, Centre National de la Recherche Scientifique (CNRS), Inserm, Université Clermont Auvergne, Clermont-Ferrand, France.
Life Sci Alliance. 2018 Dec 12;1(6):e201800197. doi: 10.26508/lsa.201800197. eCollection 2018 Dec.
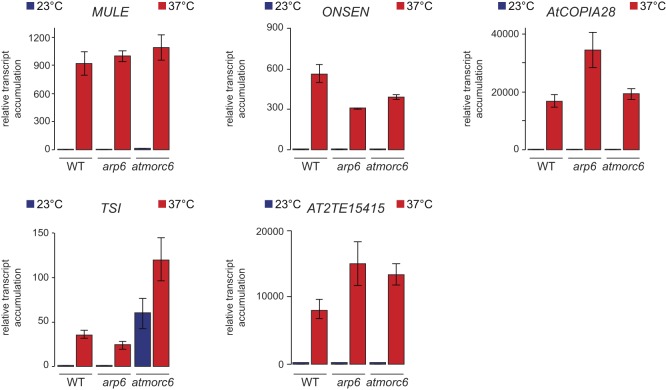
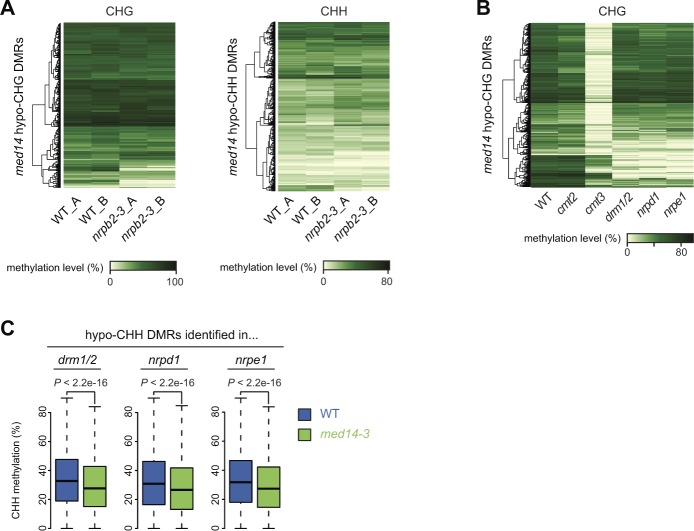
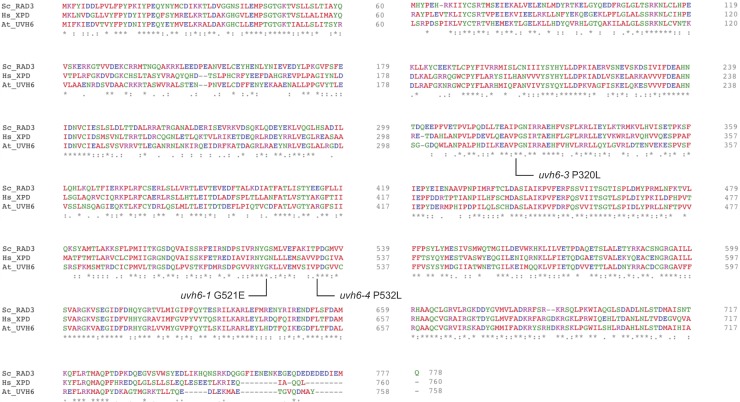
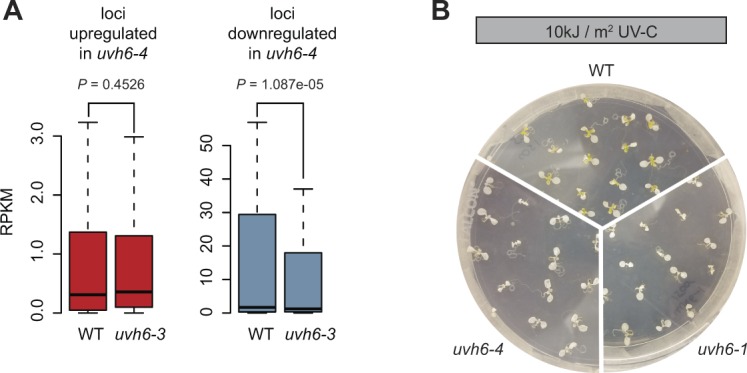
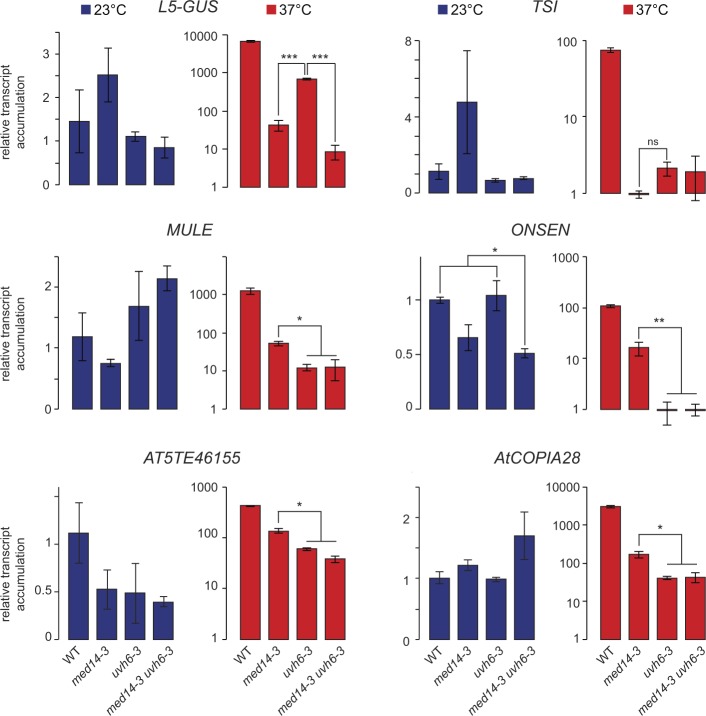
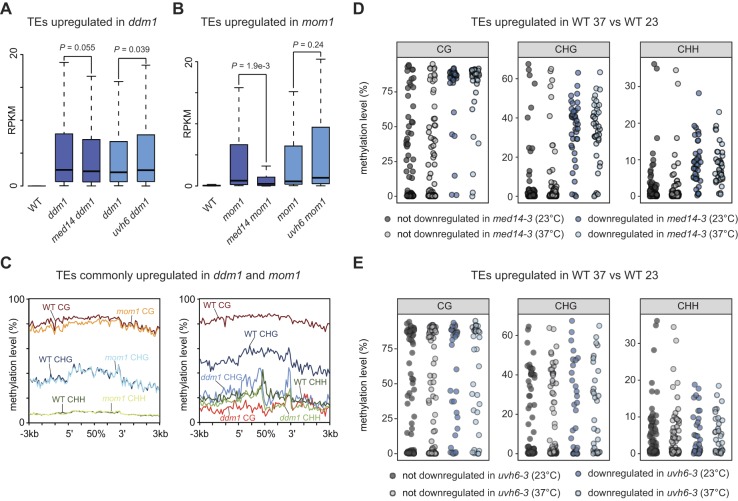
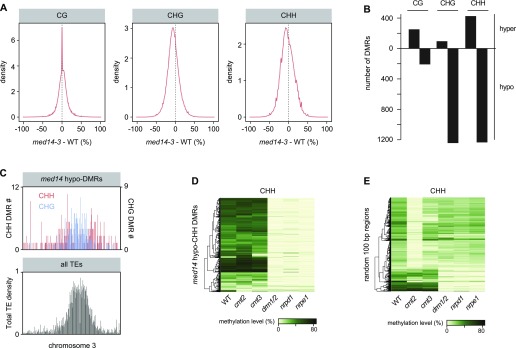
Constitutive heterochromatin is associated with repressive epigenetic modifications of histones and DNA which silence transcription. Yet, particular mutations or environmental changes can destabilize heterochromatin-associated silencing without noticeable changes in repressive epigenetic marks. Factors allowing transcription in this nonpermissive chromatin context remain poorly known. Here, we show that the transcription factor IIH component UVH6 and the mediator subunit MED14 are both required for heat stress-induced transcriptional changes and release of heterochromatin transcriptional silencing in . We find that MED14, but not UVH6, is required for transcription when heterochromatin silencing is destabilized in the absence of stress through mutating the MOM1 silencing factor. In this case, our results raise the possibility that transcription dependency over MED14 might require intact patterns of repressive epigenetic marks. We also uncover that MED14 regulates DNA methylation in non-CG contexts at a subset of RNA-directed DNA methylation target loci. These findings provide insight into the control of heterochromatin transcription upon silencing destabilization and identify MED14 as a regulator of DNA methylation.
组成型异染色质与组蛋白和DNA的抑制性表观遗传修饰相关,这些修饰会使转录沉默。然而,特定的突变或环境变化会使异染色质相关的沉默不稳定,而抑制性表观遗传标记却没有明显变化。在这种不允许转录的染色质环境中允许转录的因素仍然知之甚少。在这里,我们表明转录因子IIH成分UVH6和中介亚基MED14都是热应激诱导的转录变化和异染色质转录沉默释放所必需的。我们发现,当通过突变MOM1沉默因子在无应激情况下使异染色质沉默不稳定时,MED14而非UVH6是转录所必需的。在这种情况下,我们的结果提出了一种可能性,即对MED14的转录依赖性可能需要完整的抑制性表观遗传标记模式。我们还发现,MED14在RNA指导的DNA甲基化靶位点的一个子集中调节非CG背景下的DNA甲基化。这些发现为沉默不稳定时异染色质转录的控制提供了见解,并将MED14鉴定为DNA甲基化的调节因子。