Naturalis Biodiversity Center, RA Leiden, the Netherlands.
PLoS One. 2018 Dec 26;13(12):e0208158. doi: 10.1371/journal.pone.0208158. eCollection 2018.
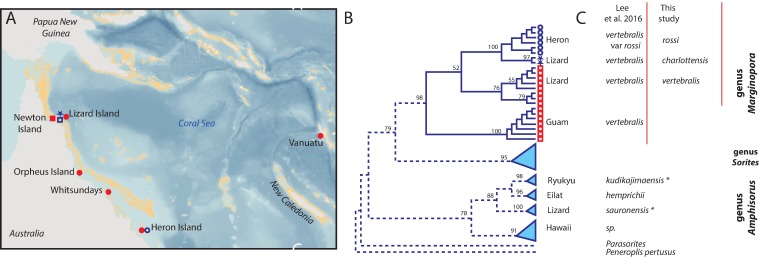
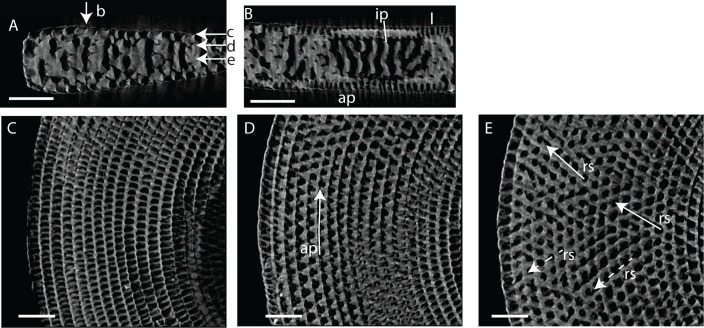
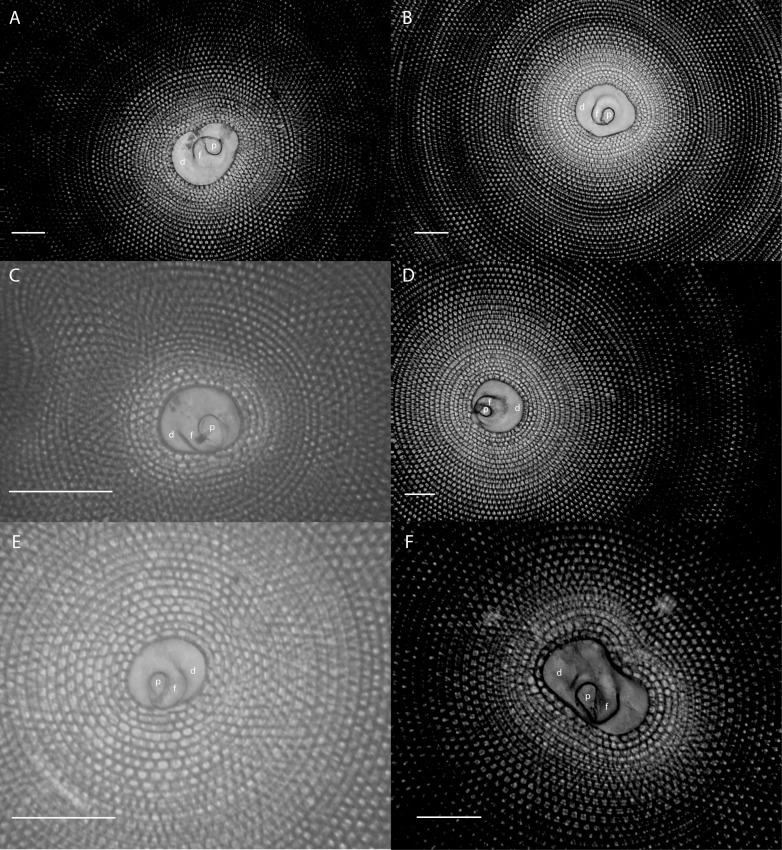
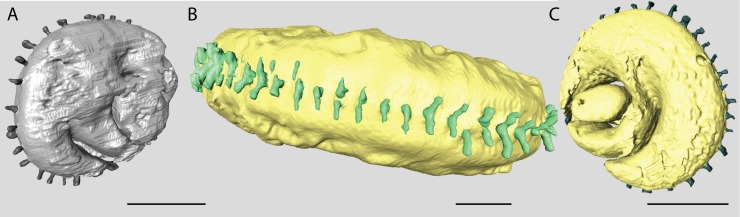
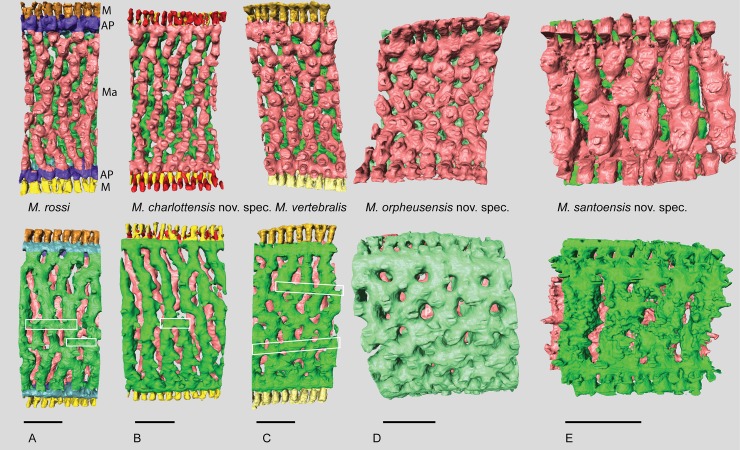
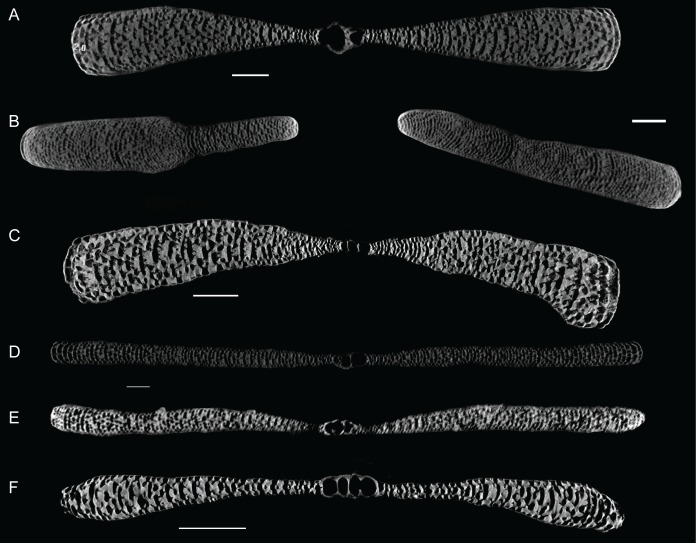
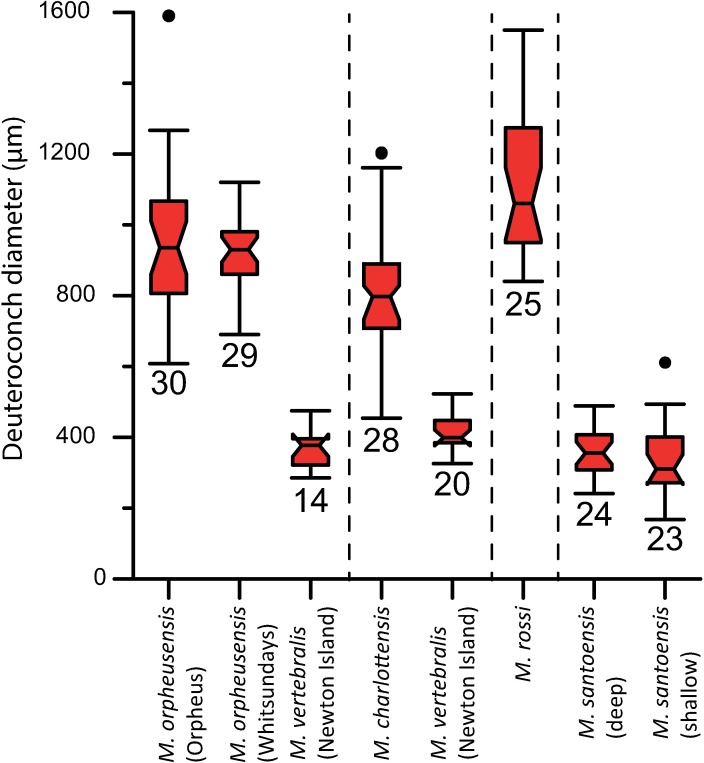
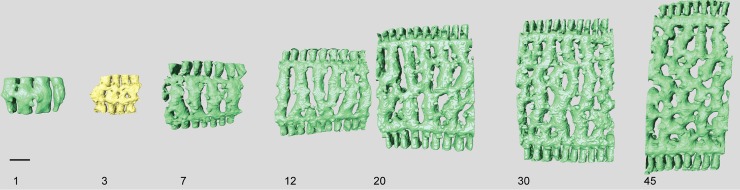
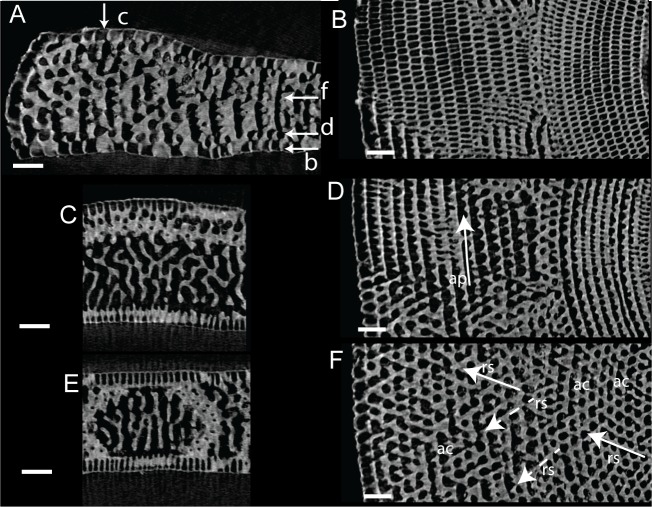
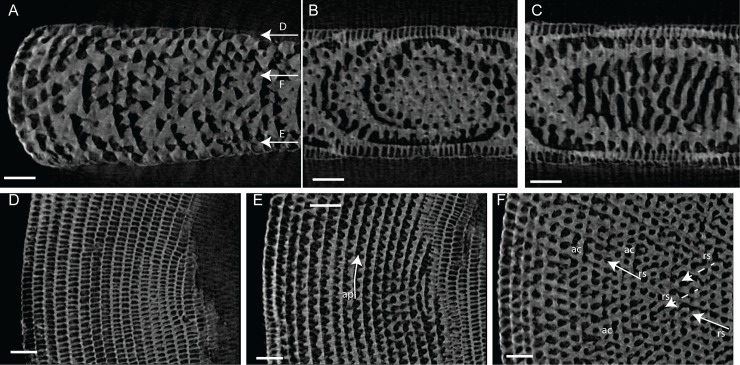
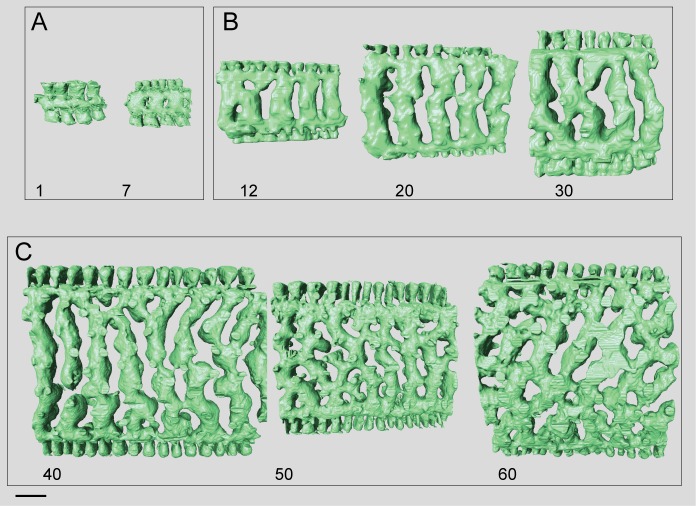
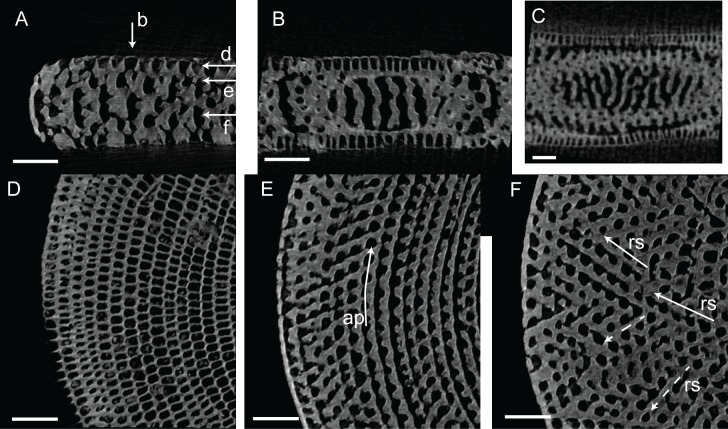
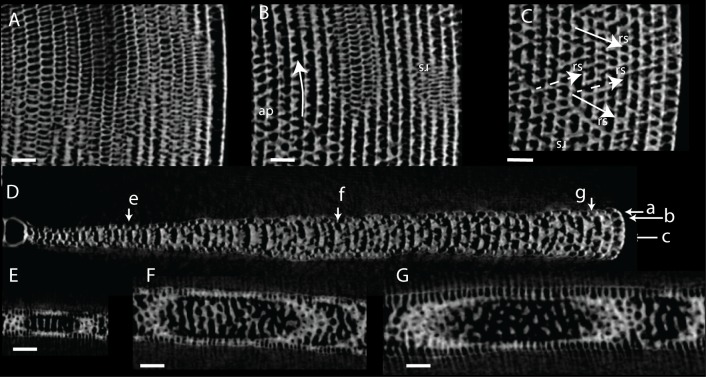
Benthic foraminifera, and certainly symbiont-bearing (large) benthic foraminifera are generally considered to have large geographic ranges in combination with significant ecomorphological variation. With the advance of molecular phylogenetic approaches, supported or preceded by detailed morphological studies, it was demonstrated that this view needs to be reevaluated. In this paper I evaluate the morphology of five Marginopora populations from around the Coral Sea by microCT-scanning. I argue that ecomorphological and ontogenetic variation is smaller than geographic variation in morphology. This forms the basis for the description of three new species, M. santoensis nov. spec., M. charlottensis nov. spec., M. orpheusensis nov. spec. Quantitative morphological variation between M. rossi, M. orpheusensis nov. spec. and M. charlottensis nov. spec. is overlapping, but each species has unique morphological characters supporting recognition as new species. Support to distinguish the deep living (M. rossi, M. charlottensis nov. spec., M. orpheusensis nov. spec.) and shallow living (M. vertebralis) Marginopora populations as separate species is strong, but not enough molecular phylogenetic data are available to test the three new deep-living species on the Great Barrier Reef hypothesis. However, detailed understanding of ecophenotypic variation in M. santoensis nov. spec. supports the conclusion that it is unlikely that ecophenotypic variation can explain the morphological variation between the three species. I argue that the number of species in this genus is underestimated, and that there are at least five species in the Coral Sea area alone.
底栖有孔虫,尤其是共生(大型)底栖有孔虫通常被认为具有广泛的地理分布范围,并伴随着显著的生态形态变异。随着分子系统发育方法的进步,在详细的形态学研究的支持或之前,已经证明这种观点需要重新评估。在本文中,我通过微 CT 扫描评估了来自珊瑚海周围的五个 Marginopora 种群的形态。我认为生态形态和个体发育变异小于形态的地理变异。这为描述三个新物种提供了基础,即 M. santoensis nov. spec.、M. charlottensis nov. spec. 和 M. orpheusensis nov. spec.。M. rossi、M. orpheusensis nov. spec. 和 M. charlottensis nov. spec. 之间的定量形态变异是重叠的,但每个物种都具有独特的形态特征,支持将其识别为新物种。区分深海生活(M. rossi、M. charlottensis nov. spec.、M. orpheusensis nov. spec.)和浅海生活(M. vertebralis) Marginopora 种群的支持是强有力的,但没有足够的分子系统发育数据来测试大堡礁假说中的三个新深海物种。然而,对 M. santoensis nov. spec. 的生态表型变异的详细了解支持这样的结论,即生态表型变异不太可能解释这三个物种之间的形态变异。我认为这个属的物种数量被低估了,仅在珊瑚海地区就至少有五个物种。