Maier Dieter
Institute of Genetics (240), University of Hohenheim, Garbenstr. 30, 70599 Stuttgart, Germany.
Hereditas. 2019 Jan 17;156:5. doi: 10.1186/s41065-019-0081-0. eCollection 2019.
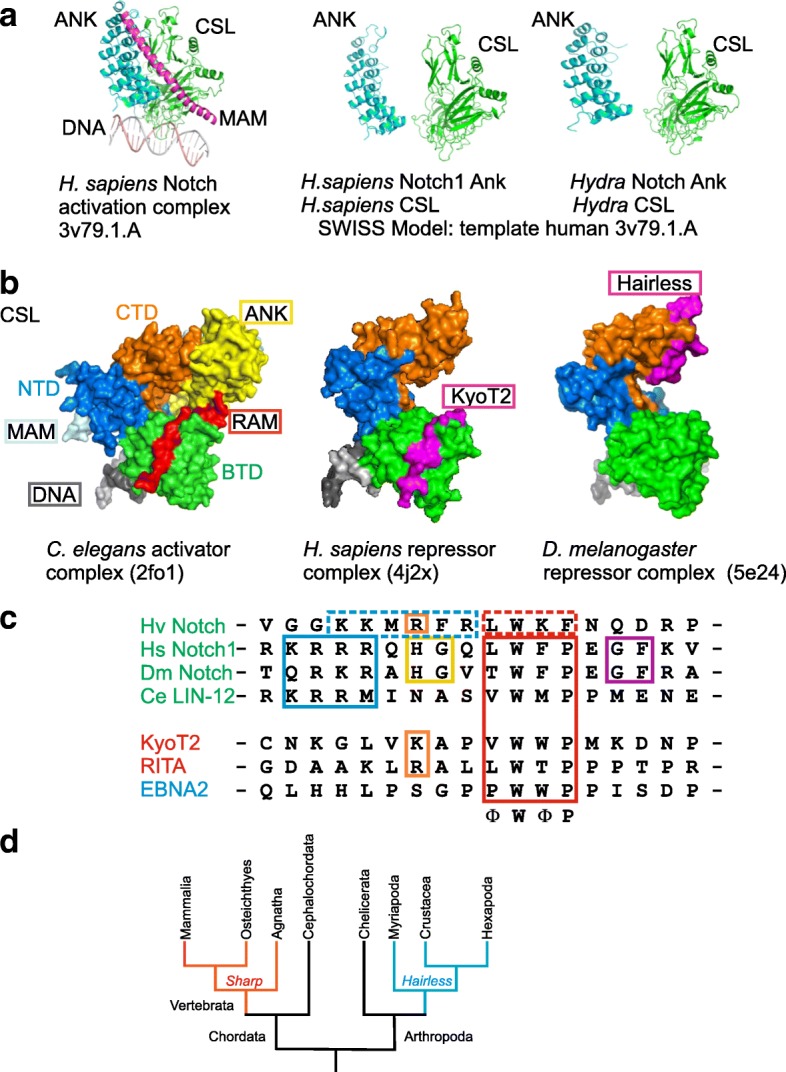
The Notch signaling pathway governs the specification of different cell types in flies, nematodes and vertebrates alike. Principal components of the pathway that activate Notch target genes are highly conserved throughout the animal kingdom. Despite the impact on development and disease, repression mechanisms are less well studied. Repressors are known from arthropods and vertebrates that differ strikingly by mode of action: whereas Hairless assembles repressor complexes with CSL transcription factors, competition between activator and repressors occurs in vertebrates (for example SHARP/MINT and KyoT2). This divergence raises questions on the evolution: Are there common ancestors throughout the animal kingdom?
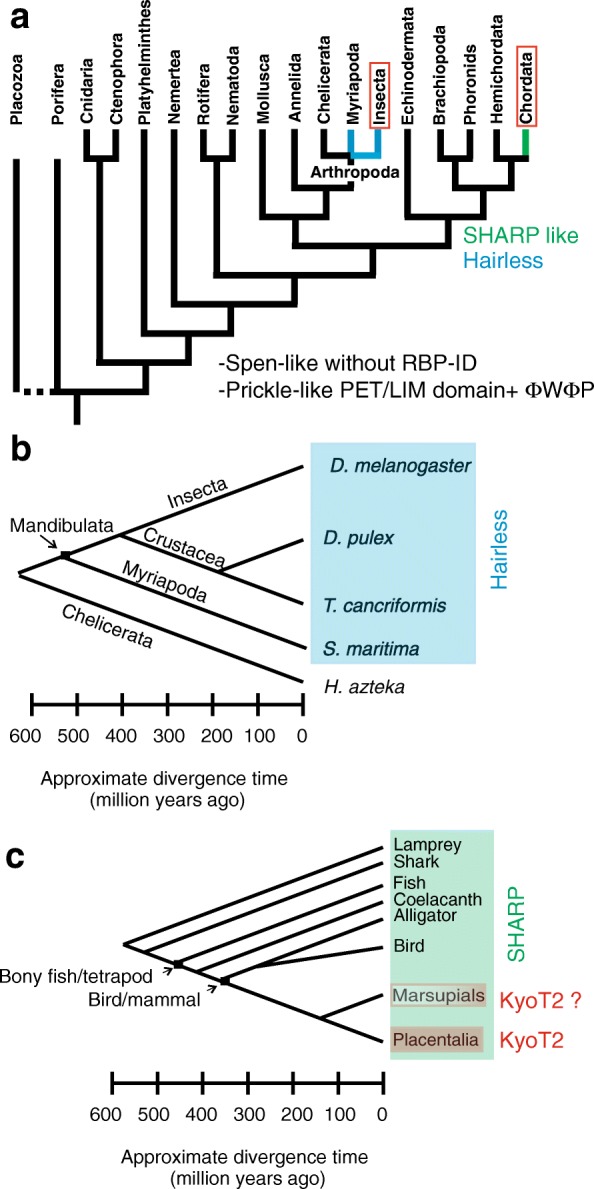
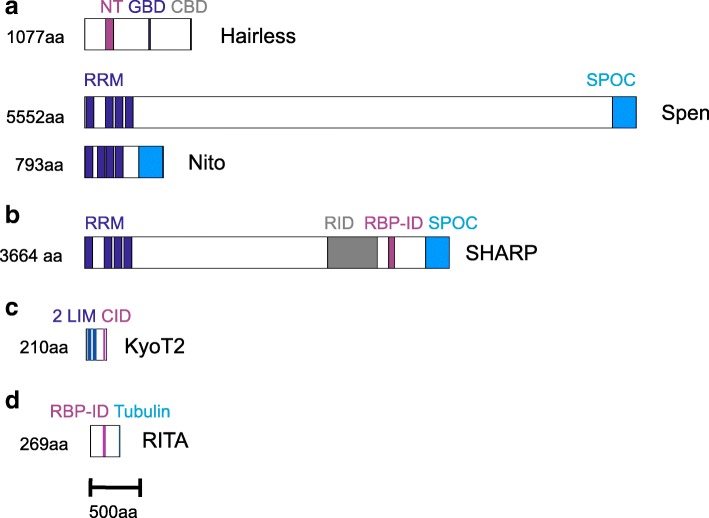
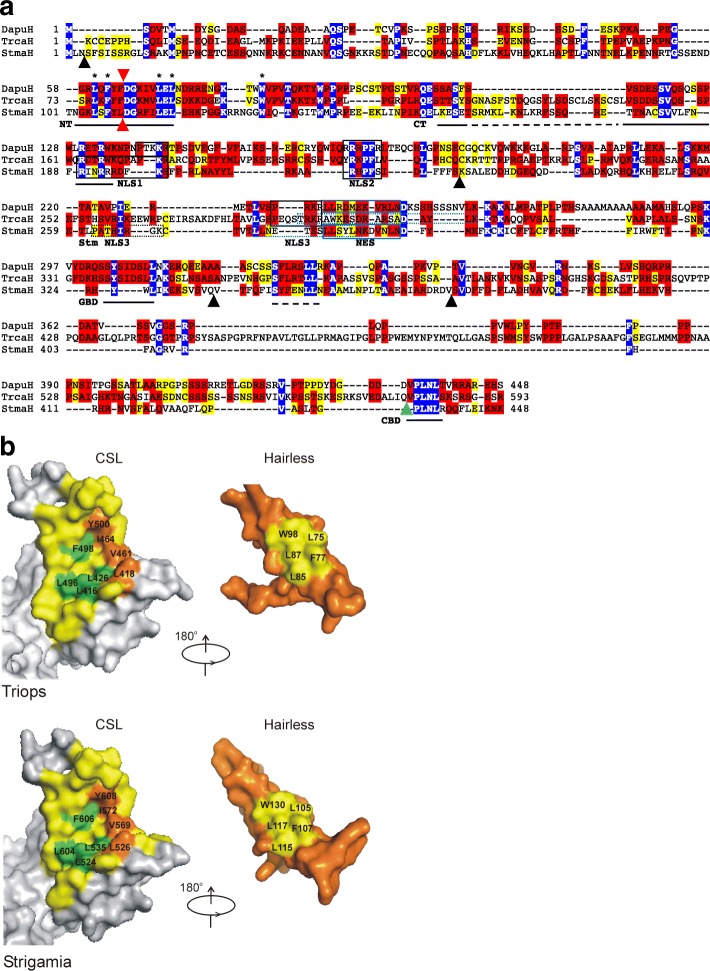
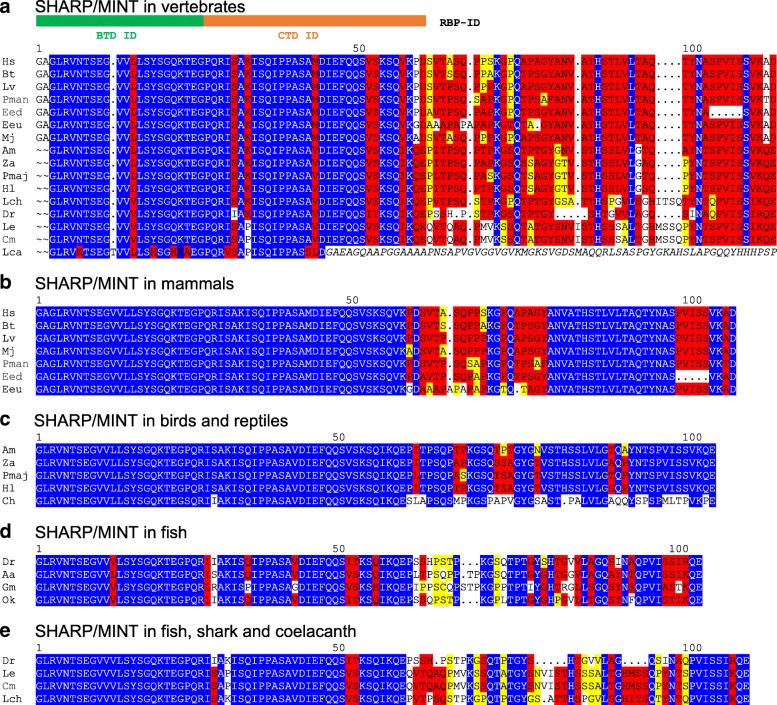
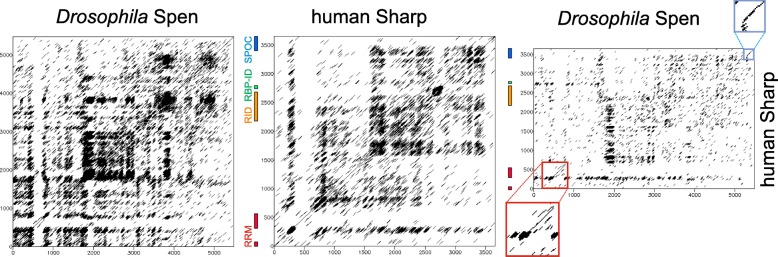
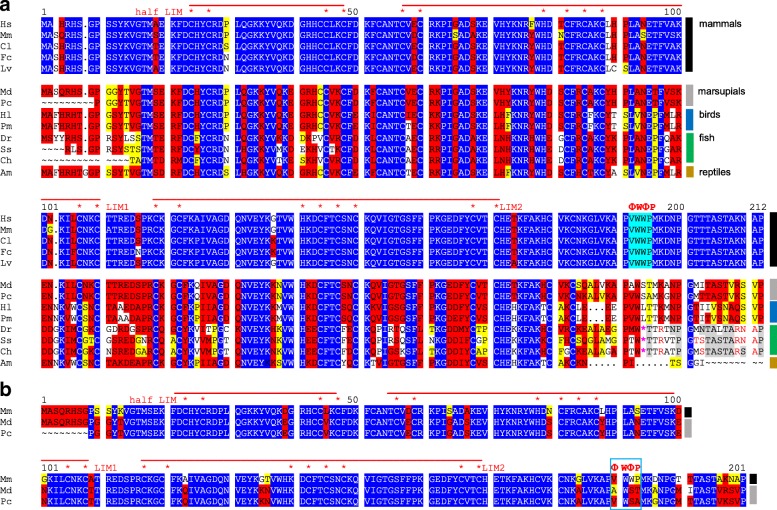
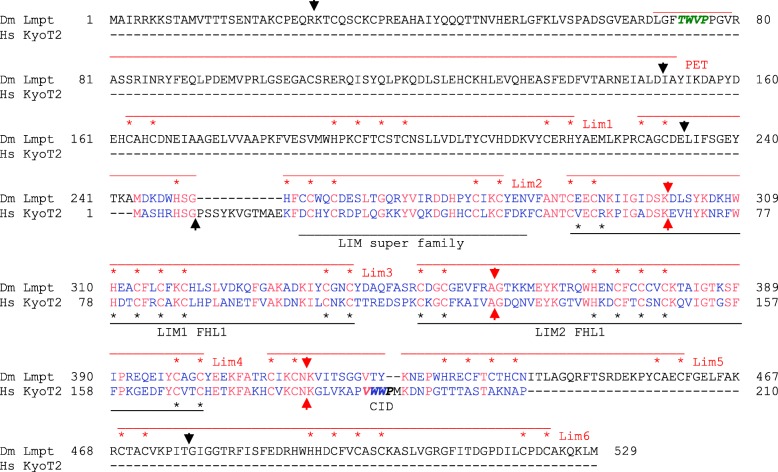
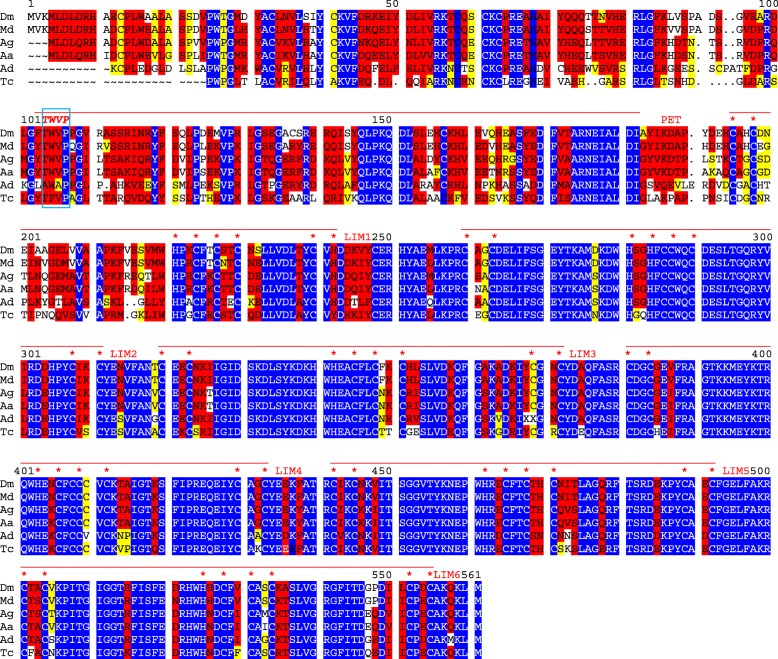
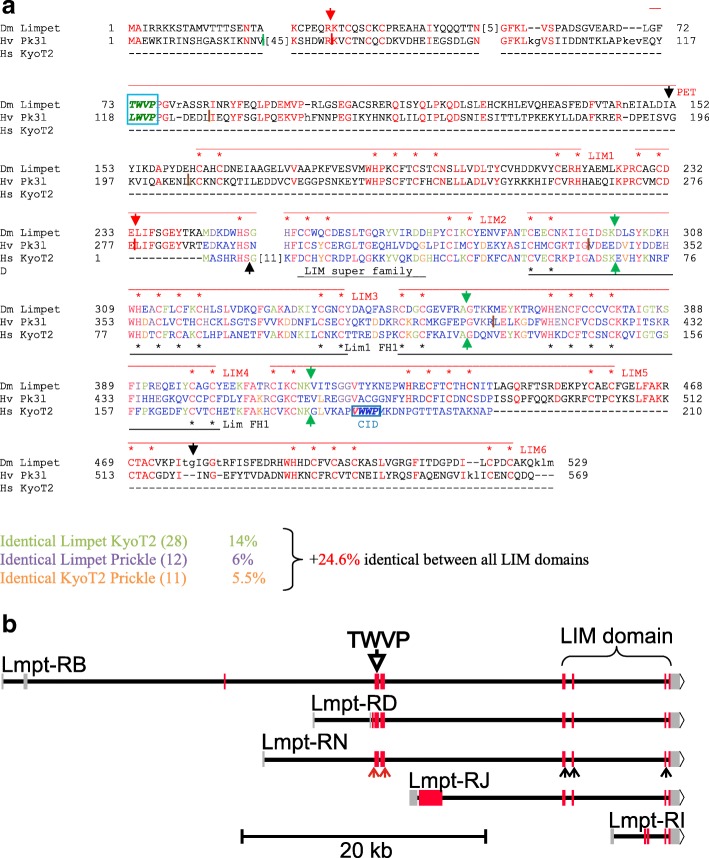
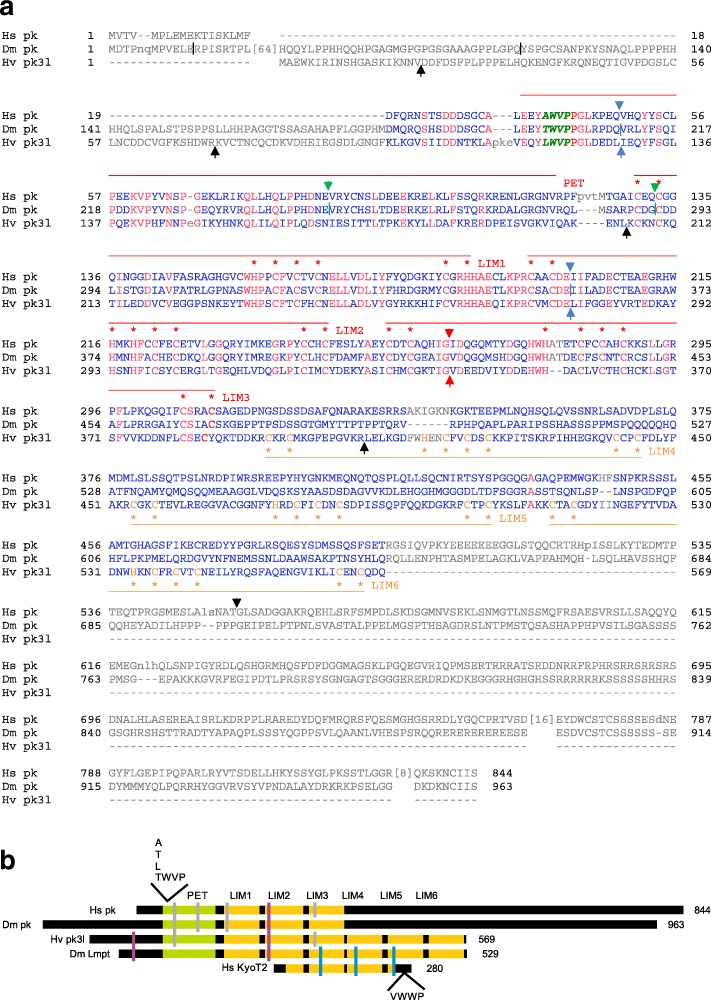
Available genome databases representing all animal clades were searched for homologues of Hairless, SHARP and KyoT2. The most distant species with convincing Hairless orthologs belong to Myriapoda, indicating its emergence after the Mandibulata-Chelicarata radiation about 500 million years ago. SHARP shares motifs with SPEN and SPENITO proteins, present throughout the animal kingdom. The CSL interacting domain of SHARP, however, is specific to vertebrates separated by roughly 600 million years of evolution. KyoT2 bears a C-terminal CSL interaction domain (CID), present only in placental mammals but highly diverged already in marsupials, suggesting introduction roughly 100 million years ago. Based on the LIM-domains that characterize KyoT2, homologues can be found in (Limpet) and (Prickle 3 like). These lack the CID of KyoT2, however, contain a PET and additional LIM domains. Conservation of intron/exon boundaries underscores the phylogenetic relationship between KyoT2, Limpet and Prickle. Most strikingly, Limpet and Prickle proteins carry a tetra-peptide motif resembling that of several CSL interactors. Overall, KyoT2 may have evolved from and to a Notch repressor in mammals.
Notch repressors appear to be specific to either chordates or arthropods. Orthologues of experimentally validated repressors were not found outside the phylogenetic group they have been originally identified. However, the data provide a hypothesis on the evolution of mammalian KyoT2 from Prickle like ancestors. The finding of a potential CSL interacting domain in Prickle homologues points to a novel, very ancestral CSL interactor present in the entire animal kingdom.
Notch信号通路在果蝇、线虫和脊椎动物中均调控着不同细胞类型的特化。该信号通路中激活Notch靶基因的主要成分在整个动物界高度保守。尽管其对发育和疾病有影响,但抑制机制的研究较少。已知节肢动物和脊椎动物中的抑制因子在作用方式上有显著差异:无翅蛋白与CSL转录因子组装成抑制复合物,而在脊椎动物中则存在激活因子与抑制因子之间的竞争(例如SHARP/MINT和KyoT2)。这种差异引发了关于进化的问题:整个动物界是否存在共同祖先?
在代表所有动物类群的可用基因组数据库中搜索无翅蛋白、SHARP和KyoT2的同源物。具有令人信服的无翅蛋白直系同源物的最远古物种属于多足纲,这表明它在约5亿年前的有颚类-螯肢类辐射之后出现。SHARP与整个动物界都存在的SPEN和SPENITO蛋白共享基序。然而,SHARP的CSL相互作用结构域是脊椎动物特有的,其进化时间约为6亿年。KyoT2具有一个C端CSL相互作用结构域(CID),仅存在于胎盘哺乳动物中,但在有袋类动物中已经高度分化,这表明它大约在1亿年前出现。基于KyoT2特有的LIM结构域,在笠贝和类似Prickle 3的蛋白中可以找到同源物。然而,它们缺乏KyoT2的CID,但包含一个PET和额外的LIM结构域。内含子/外显子边界的保守性强调了KyoT2、笠贝和Prickle之间的系统发育关系。最引人注目的是,笠贝和Prickle蛋白带有一个四肽基序,类似于几种CSL相互作用因子的基序。总体而言,KyoT2可能是从笠贝和类似Prickle的蛋白进化而来,成为哺乳动物中的一种Notch抑制因子。
Notch抑制因子似乎要么是脊索动物特有的,要么是节肢动物特有的。在最初鉴定它们的系统发育类群之外,未发现经过实验验证的抑制因子的直系同源物。然而,这些数据提供了一个关于哺乳动物KyoT2从类似Prickle的祖先进化而来的假说。在Prickle同源物中发现潜在的CSL相互作用结构域,表明在整个动物界存在一种新的、非常古老的CSL相互作用因子。