Department of Biosciences, COMSATS University Islamabad, Islamabad, Pakistan.
Department of Biology, University of Padova, Padova, Italy.
PLoS One. 2019 Apr 30;14(4):e0213986. doi: 10.1371/journal.pone.0213986. eCollection 2019.
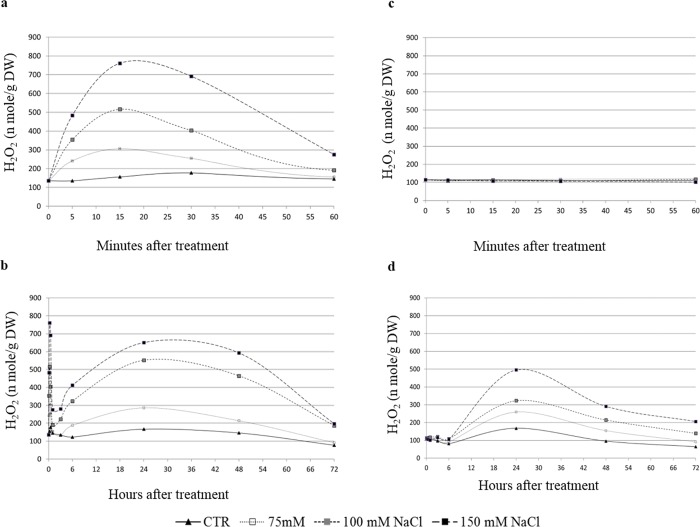
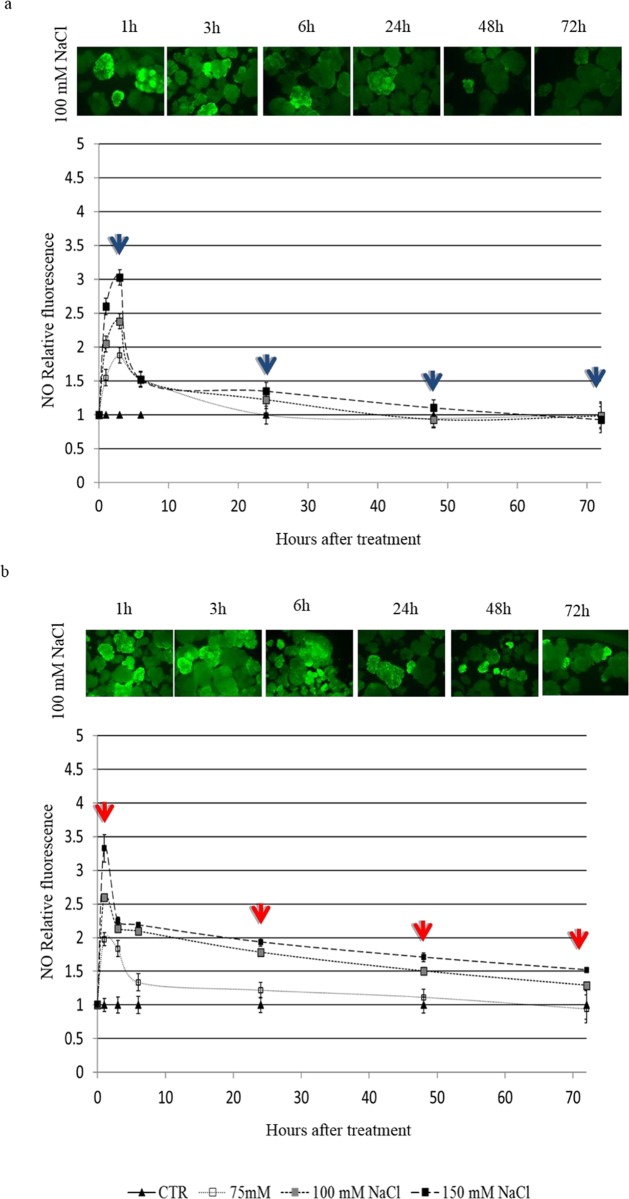
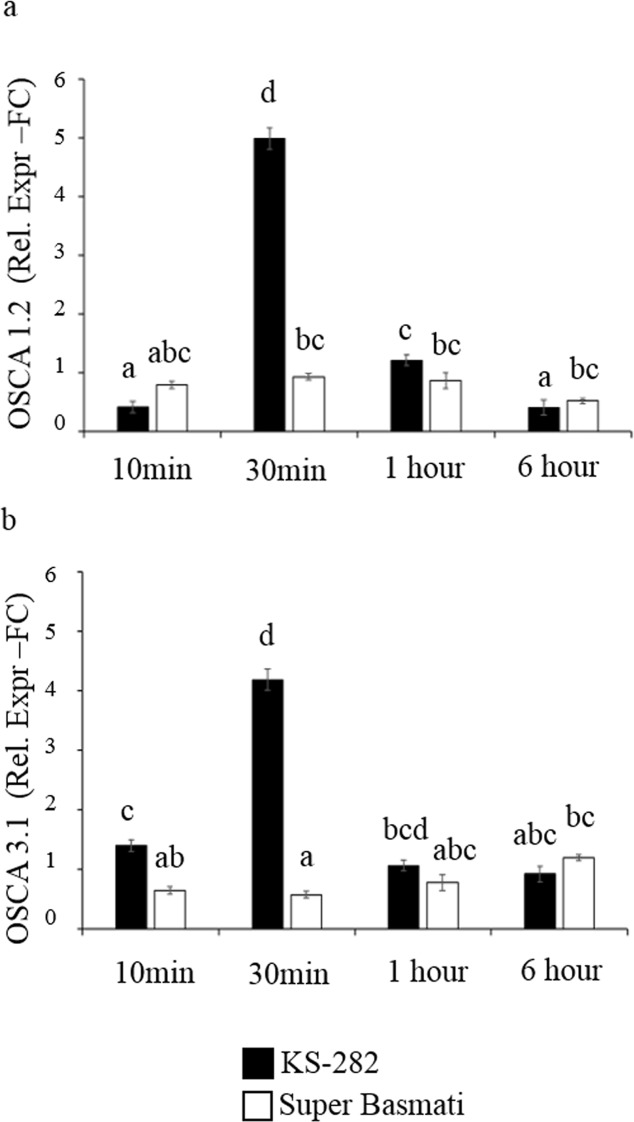
Among cereal crops, salinity tolerance is rare and complex. Multiple genes control numerous pathways, which constitute plant's response to salinity. Cell cultures act as model system and are useful to investigate the salinity response which can possibly mimic a plant's response to stress. In the present study two indica rice varieties, KS-282 and Super Basmati which exhibited contrasting sodium chloride (NaCl) stress response were used to establish cell cultures. The cell cultures showed a contrasting response to salt stress at 100 mM NaCl. High level of intracellular hydrogen peroxide (H2O2) and nitric oxide (NO) were observed in sensitive cell culture for prolonged period as compared to the tolerant cells in which an extracellular H2O2 burst along with controlled intracellular H2O2 and NO signal was seen. To evaluate the role of NO in inducing cell death under salt stress, cell death percentage (CDP) was measured after 2-4-carboxyphenyl-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (cPTIO) pre-treatment. CDP was reduced significantly in both tolerant and sensitive cell cultures emphasizing NO's possible role in programmed cell death. Expression analysis of apoplastic NADPH oxidase, i.e. OsRbohA and recently characterised OSCA family members i.e. OsOSCA 1.2 and OsOSCA 3.1 was done. Intracellular H2O2/NO levels displayed an interplay between Ca2+ influx and ROS/RNS signal. Detoxifying enzyme (i.e. ascorbate peroxidase and catalase) activity was considerably higher in tolerant KS-282 while the activity of superoxide dismutase was significantly prominent in the sensitive cells triggering greater oxidative damage owing to the prolonged presence of intracellular H2O2. Salt stress and ROS responsive TFs i.e. OsSERF1 and OsDREB2A were expressed exclusively in the tolerant cells. Similarly, the expression of genes involved in maintaining high [K+]/[Na+] ratio was considerably higher and earlier in the tolerant variety. Overall, we suggest that a control over ROS production, and an increase in the expression of genes important for potassium homeostasis play a dynamic role in salinity tolerance in rice cell cultures.
在谷类作物中,耐盐性是罕见且复杂的。多个基因控制着众多途径,这些途径构成了植物对盐度的响应。细胞培养作为一种模型系统,可用于研究盐度响应,该响应可能模拟植物对胁迫的响应。在本研究中,使用了两种耐盐性不同的籼稻品种 KS-282 和 Super Basmati 来建立细胞培养物。细胞培养物在 100mM NaCl 胁迫下表现出不同的响应。与耐盐细胞相比,敏感细胞培养物中观察到高水平的细胞内过氧化氢(H2O2)和一氧化氮(NO),并且在较长时间内持续存在。在耐盐细胞中,观察到细胞外 H2O2 爆发以及受控的细胞内 H2O2 和 NO 信号。为了评估 NO 在盐胁迫下诱导细胞死亡的作用,在用 2-4-羧基苯基-4,4,5,5-四甲基咪唑啉-1-氧-3-氧化物(cPTIO)预处理后测量细胞死亡百分比(CDP)。在耐盐和敏感细胞培养物中,CDP 均显著降低,这强调了 NO 在程序性细胞死亡中的可能作用。对质外体 NADPH 氧化酶,即 OsRbohA 和最近鉴定的 OSCA 家族成员,即 OsOSCA 1.2 和 OsOSCA 3.1 的表达进行了分析。细胞内 H2O2/NO 水平显示 Ca2+流入和 ROS/RNS 信号之间的相互作用。在耐盐的 KS-282 中,解毒酶(即抗坏血酸过氧化物酶和过氧化氢酶)的活性明显较高,而在敏感细胞中,超氧化物歧化酶的活性非常明显,这导致由于细胞内 H2O2 的长时间存在而引起更大的氧化损伤。盐胁迫和 ROS 响应转录因子,即 OsSERF1 和 OsDREB2A,仅在耐盐细胞中表达。同样,维持高[K+]/[Na+]比的基因的表达在耐盐品种中也更早更高。总的来说,我们认为对 ROS 产生的控制以及对钾稳态重要基因的表达增加在水稻细胞培养物的耐盐性中起着动态作用。