Ministry of Education Key Laboratory of Biosystems Homeostasis & Protection and Innovation Center for Cell Signaling Network, Life Sciences Institute, Zhejiang University, 310058 Hangzhou, Zhejiang, China.
Ministry of Education Key Laboratory of Biosystems Homeostasis & Protection and Innovation Center for Cell Signaling Network, Life Sciences Institute, Zhejiang University, 310058 Hangzhou, Zhejiang, China
Proc Natl Acad Sci U S A. 2019 Oct 22;116(43):21580-21591. doi: 10.1073/pnas.1911745116. Epub 2019 Oct 9.
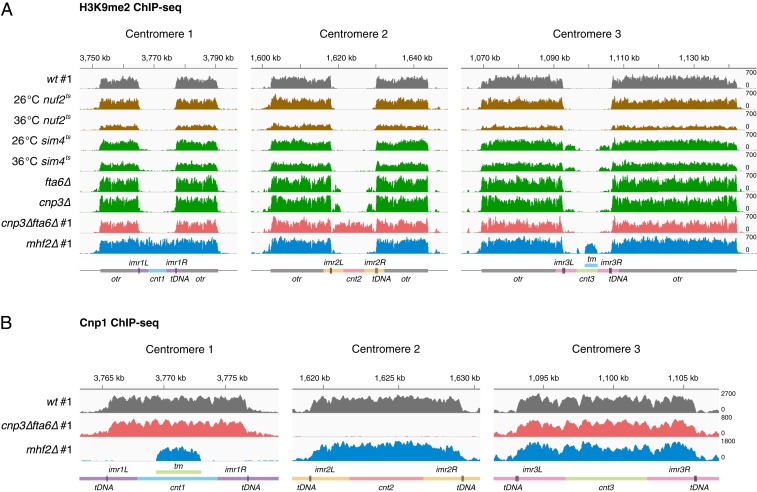
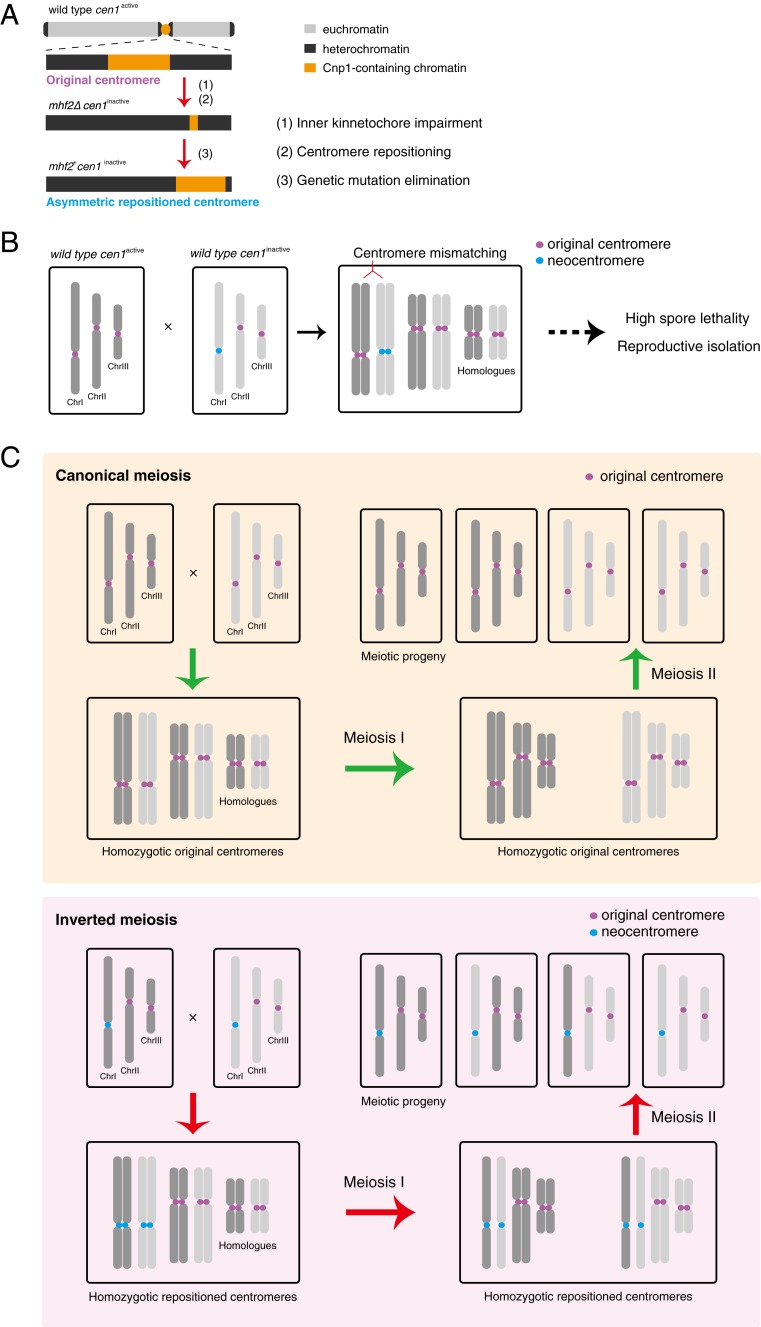
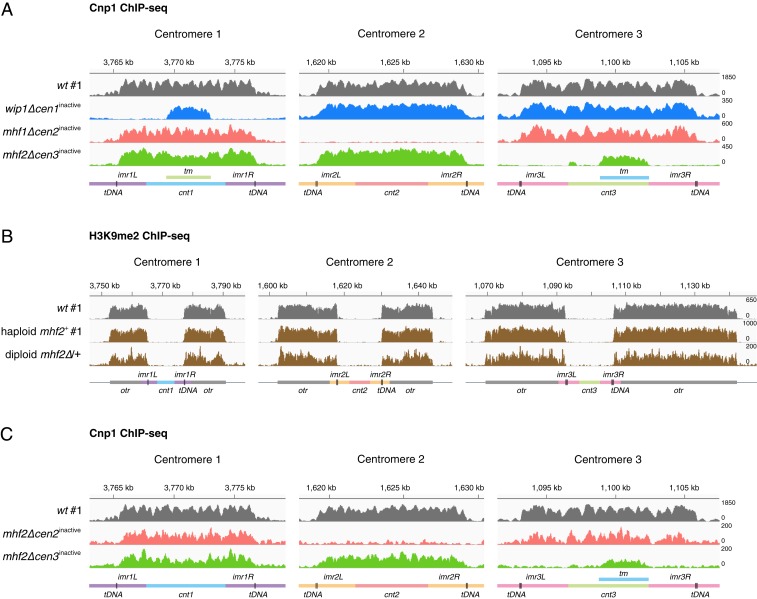
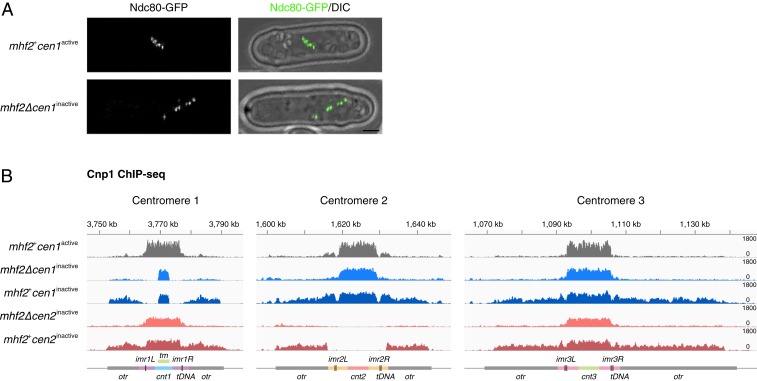
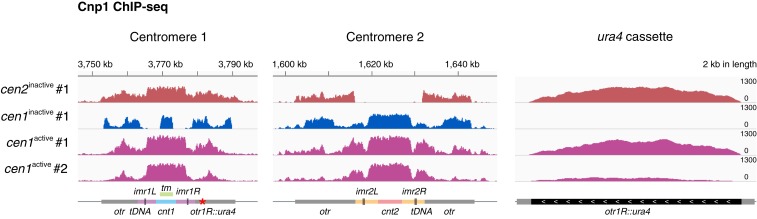
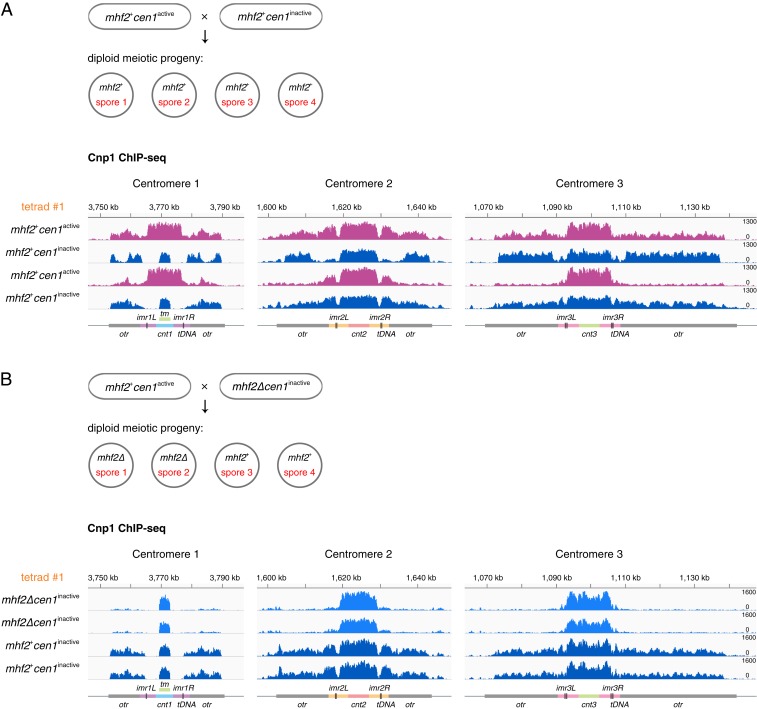
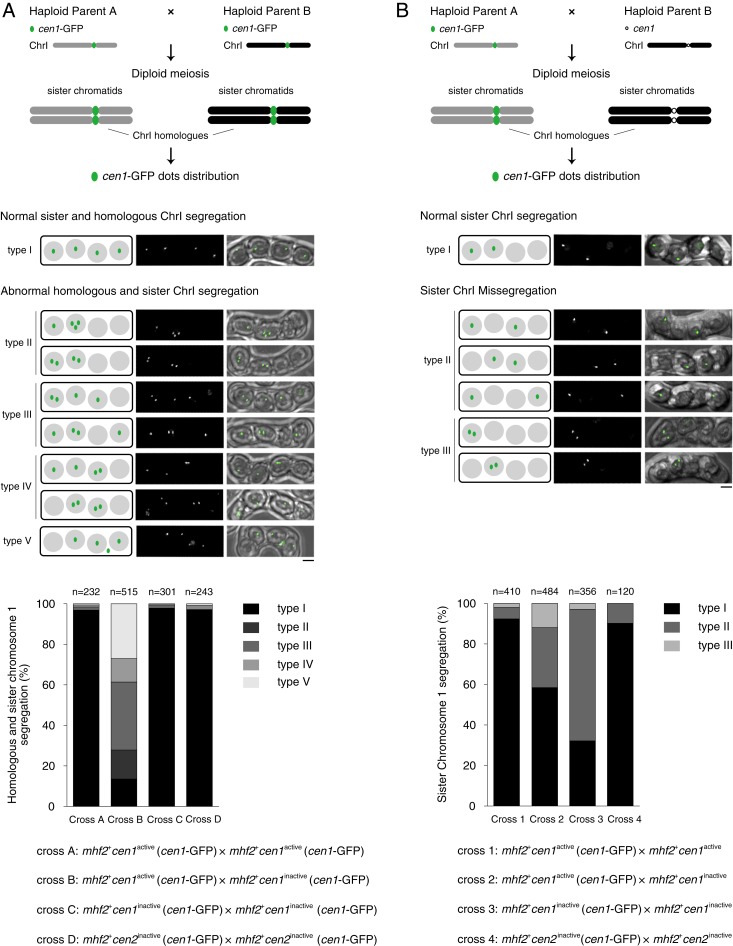
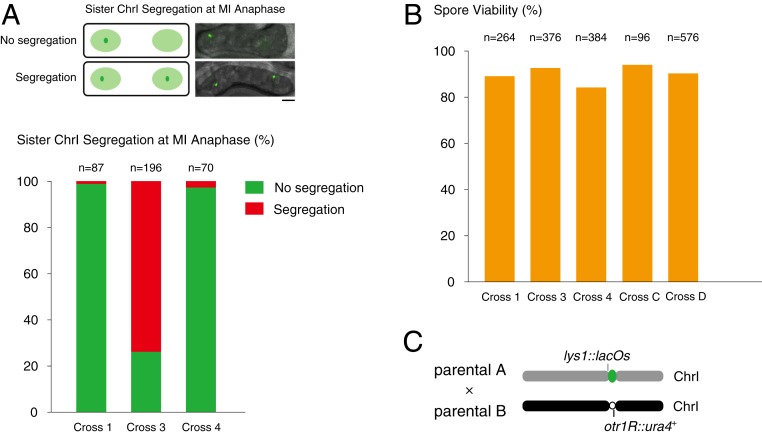
The chromosomal position of each centromere is determined epigenetically and is highly stable, whereas incremental cases have supported the occurrence of centromere repositioning on an evolutionary time scale (evolutionary new centromeres, ENCs), which is thought to be important in speciation. The mechanisms underlying the high stability of centromeres and its functional significance largely remain an enigma. Here, in the fission yeast , we identify a feedback mechanism: The kinetochore, whose assembly is guided by the centromere, in turn, enforces centromere stability. Upon going through meiosis, specific inner kinetochore mutations induce centromere repositioning-inactivation of the original centromere and formation of a new centromere elsewhere-in 1 of the 3 chromosomes at random. Repositioned centromeres reside asymmetrically in the pericentromeric regions and cells carrying them are competent in mitosis and homozygotic meiosis. However, when cells carrying a repositioned centromere are crossed with those carrying the original centromere, the progeny suffer severe lethality due to defects in meiotic chromosome segregation. Thus, repositioned centromeres constitute a reproductive barrier that could initiate genetic divergence between 2 populations with mismatched centromeres, documenting a functional role of ENCs in speciation. Surprisingly, homozygotic repositioned centromeres tend to undergo meiosis in an inverted order-that is, sister chromatids segregate first, and homologous chromosomes separate second-whereas the original centromeres on other chromosomes in the same cell undergo meiosis in the canonical order, revealing hidden flexibility in the perceived rigid process of meiosis.
每个着丝粒的染色体位置都是通过表观遗传确定的,高度稳定,而增量案例支持着丝粒在进化时间尺度上重新定位(进化中新着丝粒,ENC),这被认为在物种形成中很重要。着丝粒高度稳定性的机制及其功能意义在很大程度上仍然是一个谜。在这里,在裂殖酵母中,我们发现了一个反馈机制:由着丝粒指导组装的动粒反过来又加强了着丝粒的稳定性。在经历减数分裂时,特定的内动粒突变会导致着丝粒重新定位——原始着丝粒失活,并在其余的 3 条染色体中的任意一条上形成新的着丝粒。重新定位的着丝粒不对称地位于着丝粒周围区域,携带它们的细胞在有丝分裂和纯合减数分裂中具有活性。然而,当携带重新定位的着丝粒的细胞与携带原始着丝粒的细胞杂交时,由于减数分裂染色体分离的缺陷,后代会遭受严重的致死性。因此,重新定位的着丝粒构成了一个生殖障碍,可能会在两个带有不匹配着丝粒的种群之间引发遗传分化,证明了 ENC 在物种形成中的功能作用。令人惊讶的是,纯合重新定位的着丝粒往往会以相反的顺序进行减数分裂——即姐妹染色单体首先分离,同源染色体随后分离——而同一细胞中其他染色体上的原始着丝粒则以经典顺序进行减数分裂,揭示了减数分裂这一被认为是僵化过程中的隐藏灵活性。