Department of Biochemistry and Molecular Biology, The University of Kansas Medical Center, Kansas City, KS, 66160, USA.
Sci Rep. 2019 Nov 18;9(1):16957. doi: 10.1038/s41598-019-53464-z.
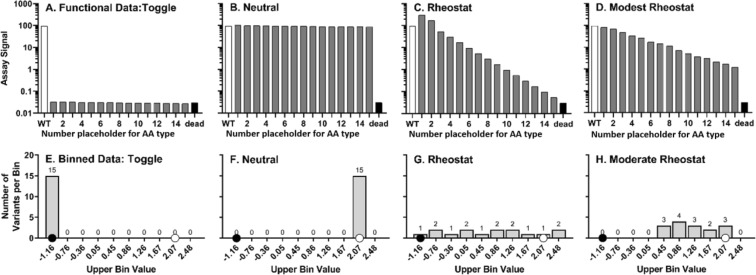
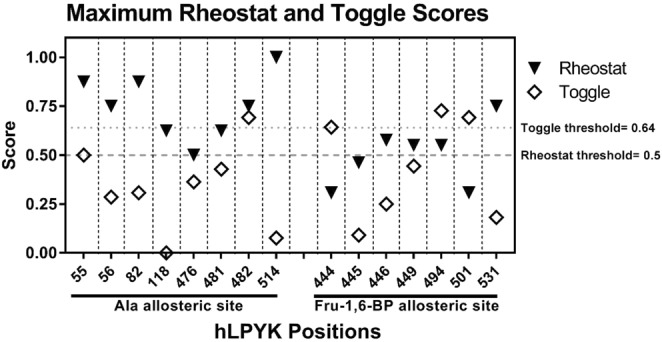
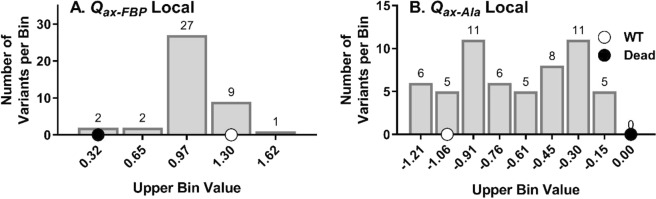
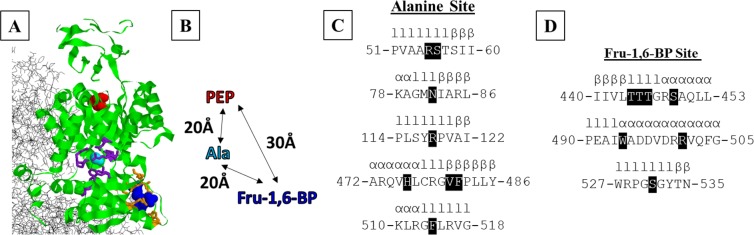
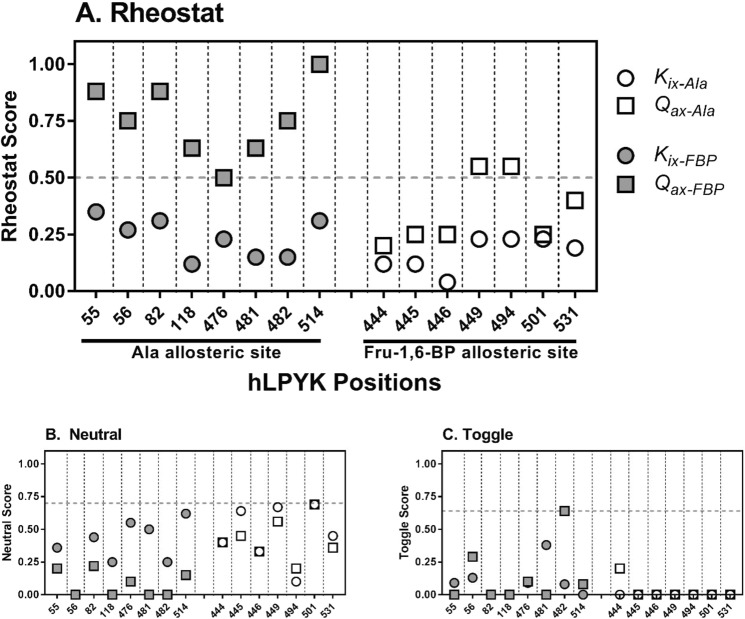
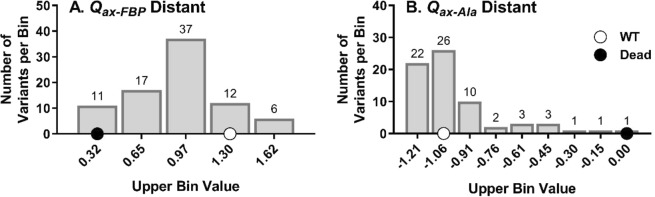
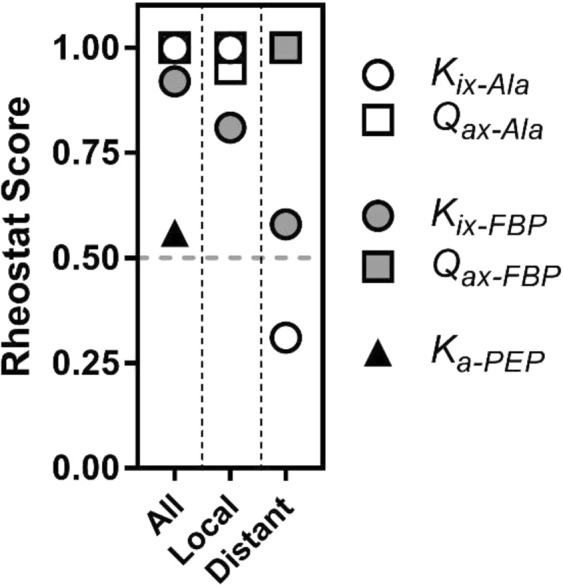
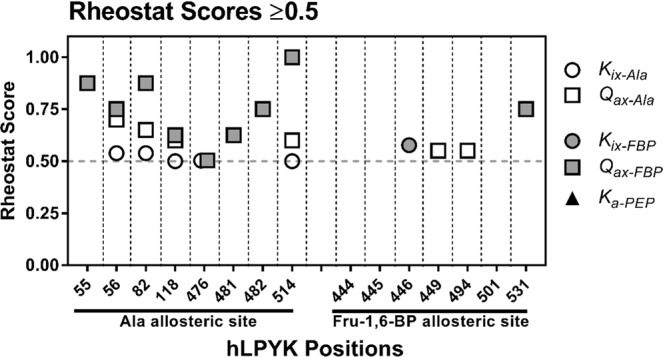
For protein mutagenesis, a common expectation is that important positions will behave like on/off "toggle" switches (i.e., a few substitutions act like wildtype, most abolish function). However, there exists another class of important positions that manifests a wide range of functional outcomes upon substitution: "rheostat" positions. Previously, we evaluated rheostat positions located near the allosteric binding sites for inhibitor alanine (Ala) and activator fructose-1,6-bisphosphate (Fru-1,6-BP) in human liver pyruvate kinase. When substituted with multiple amino acids, many positions demonstrated moderate rheostatic effects on allosteric coupling between effector binding and phosphoenolpyruvate (PEP) binding in the active site. Nonetheless, the combined outcomes of all positions sampled the full range of possible allosteric coupling (full tunability). However, that study only evaluated allosteric tunability of "local" positions, i.e., positions were located near the binding sites of the allosteric ligand being assessed. Here, we evaluated tunability of allosteric coupling when mutated sites were distant from the allosterically-coupled binding sites. Positions near the Ala binding site had rheostatic outcomes on allosteric coupling between Fru-1,6-BP and PEP binding. In contrast, positions in the Fru-1,6-BP site exhibited modest effects on coupling between Ala and PEP binding. Analyzed in aggregate, both PEP/Ala and PEP/Fru-1,6-BP coupling were again fully tunable by amino acid substitutions at this limited set of distant positions. Furthermore, some positions exhibited rheostatic control over multiple parameters and others exhibited rheostatic effects on one parameter and toggle control over a second. These findings highlight challenges in efforts to both predict/interpret mutational outcomes and engineer functions into proteins.
对于蛋白质的突变,一个常见的期望是重要的位置将表现得像开/关“拨动”开关(即,少数取代类似于野生型,大多数则消除功能)。然而,还有另一类重要的位置,在取代时会表现出广泛的功能结果:“变阻器”位置。以前,我们评估了位于人肝丙酮酸激酶的别构结合位点附近的变阻器位置,这些位置分别靠近抑制剂丙氨酸(Ala)和激活剂果糖-1,6-二磷酸(Fru-1,6-BP)的别构结合位点。当用多种氨基酸取代时,许多位置在别构效应器结合与活性位点中磷酸烯醇丙酮酸(PEP)结合之间的偶联中表现出中等程度的变阻器效应。尽管如此,所有位置的综合结果采样了所有可能的别构偶联的全部范围(全可调性)。然而,该研究仅评估了“局部”位置的别构可调性,即位置位于正在评估的别构配体的结合位点附近。在这里,我们评估了突变位置远离别构偶联结合位点时别构偶联的可调性。Ala 结合位点附近的位置对 Fru-1,6-BP 和 PEP 结合之间的别构偶联具有变阻器结果。相比之下,Fru-1,6-BP 位点的位置对 Ala 和 PEP 结合之间的偶联仅有适度的影响。总体分析,在这一组有限的远距离位置的氨基酸取代下,PEP/Ala 和 PEP/Fru-1,6-BP 偶联再次完全可调。此外,一些位置对多个参数具有变阻器控制作用,而另一些位置对一个参数具有变阻器控制作用,对第二个参数具有拨动控制作用。这些发现突出了在预测/解释突变结果和将功能工程化到蛋白质中所面临的挑战。