Department of Biochemistry, Microbiology, and Immunology, Wayne State University School of Medicine, Detroit, Michigan, United States of America.
PLoS One. 2020 Mar 3;15(3):e0229494. doi: 10.1371/journal.pone.0229494. eCollection 2020.
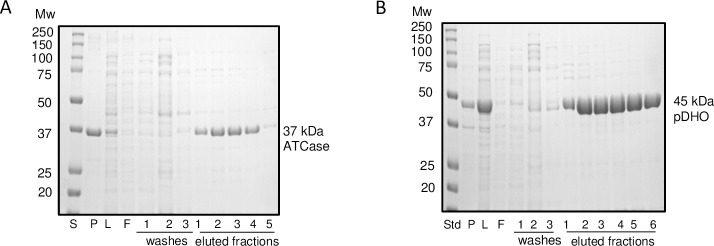
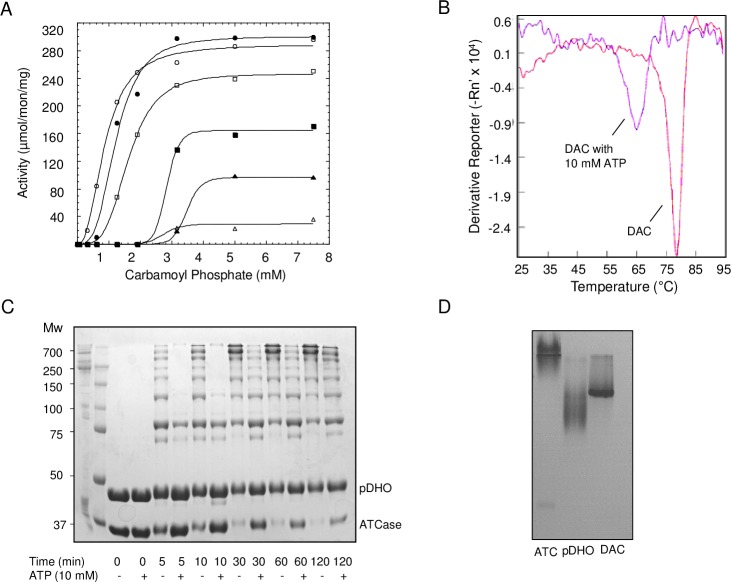
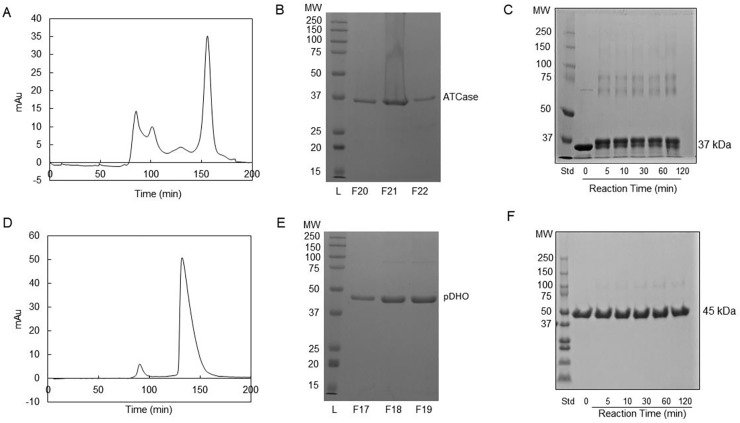
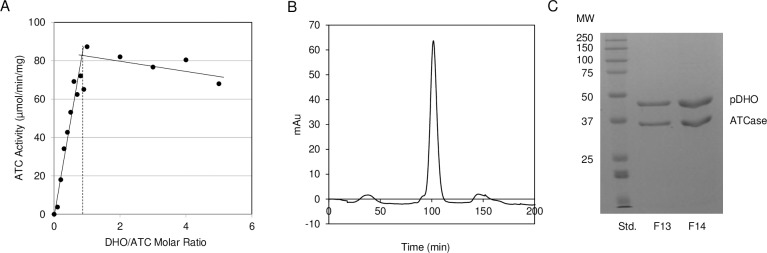
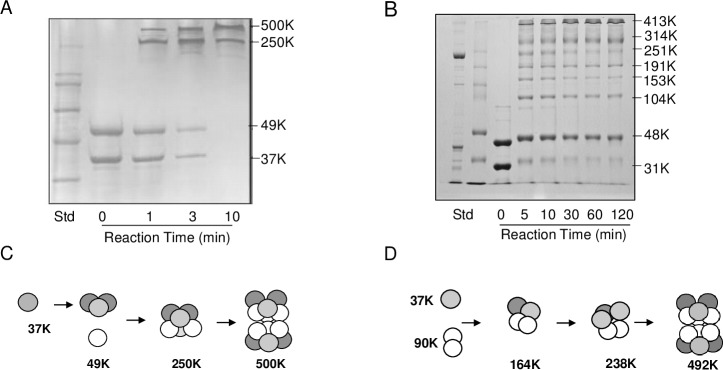
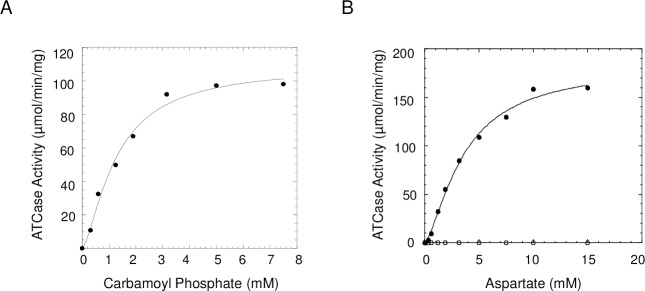
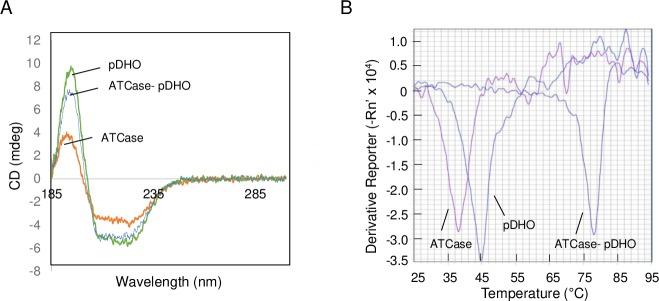
Pseudomonas aeruginosa is a virulent pathogen that has become more threatening with the emergence of multidrug resistance. The aspartate transcarbamoylase (ATCase) of this organism is a dodecamer comprised of six 37 kDa catalytic chains and six 45 kDa chains homologous to dihydroorotase (pDHO). The pDHO chain is inactive but is necessary for ATCase activity. A stoichiometric mixture of the subunits associates into a dodecamer with full ATCase activity. Unlike other known ATCases, the P. aeruginosa catalytic chain does not spontaneously assemble into a trimer. Chemical-crosslinking and size-exclusion chromatography showed that P. aeruginosa ATCase is monomeric which accounts for its lack of catalytic activity since the active site is a composite comprised of residues from adjacent monomers in the trimer. Circular dichroism spectroscopy indicated that the ATCase chain adopts a structure that contains secondary structure elements although neither the ATCase nor the pDHO subunits are very stable as determined by a thermal shift assay. Formation of the complex increases the melting temperature by about 30°C. The ATCase is strongly inhibited by all nucleotide di- and triphosphates and exhibits extreme cooperativity. Previous studies suggested that the regulatory site is located in an 11-residue extension of the amino end of the catalytic chain. However, deletion of the extensions did not affect catalytic activity, nucleotide inhibition or the assembly of the dodecamer. Nucleotides destabilized the dodecamer which probably accounts for the inhibition and apparent cooperativity of the substrate saturation curves. Contrary to previous interpretations, these results suggest that P. aeruginosa ATCase is not allosterically regulated by nucleotides.
铜绿假单胞菌是一种毒力很强的病原体,随着多药耐药性的出现,它变得更加危险。该生物体的天冬氨酸转氨甲酰酶(ATCase)是由六个 37 kDa 催化链和六个与二氢乳清酸酶(pDHO)同源的 45 kDa 链组成的十二聚体。pDHO 链无活性,但对 ATCase 活性是必需的。亚基的化学计量混合物缔合形成具有完整 ATCase 活性的十二聚体。与其他已知的 ATCases 不同,铜绿假单胞菌的催化链不会自发组装成三聚体。化学交联和大小排阻色谱显示,铜绿假单胞菌 ATCase 是单体,这解释了其缺乏催化活性,因为活性位点是由三聚体中相邻单体的残基组成的复合物。圆二色性光谱表明,ATCase 链采用一种结构,其中包含二级结构元件,尽管 ATCase 和 pDHO 亚基都不是非常稳定,如热移测定所确定的。形成复合物使熔点提高约 30°C。ATCase 强烈抑制所有核苷酸二磷酸和三磷酸,并表现出极端的协同性。先前的研究表明,调节部位位于催化链氨基末端的 11 个残基延伸处。然而,删除延伸部分并不影响催化活性、核苷酸抑制或十二聚体的组装。核苷酸使十二聚体不稳定,这可能解释了抑制和底物饱和曲线的明显协同性。与先前的解释相反,这些结果表明,铜绿假单胞菌 ATCase 不是由核苷酸变构调节的。