Department of Microbiology and Immunology, Dalhousie University, Halifax, NS B3H 4R2, Canada.
Department of Biochemistry and Molecular Biology, Dalhousie University, Halifax, NS B3H 4R2, Canada.
Viruses. 2020 Jun 29;12(7):702. doi: 10.3390/v12070702.
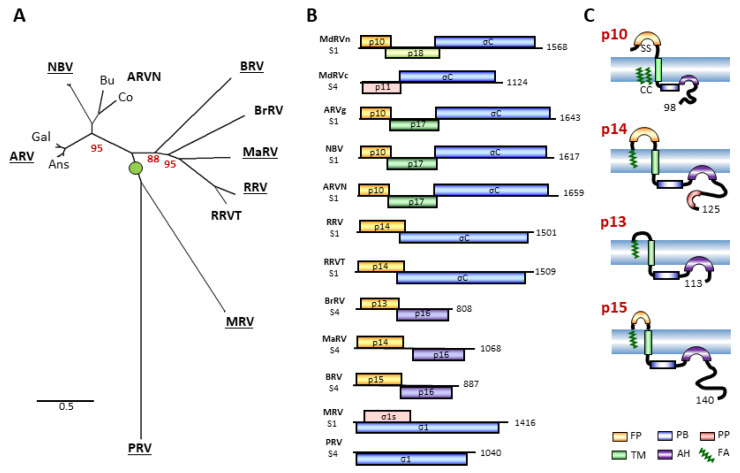
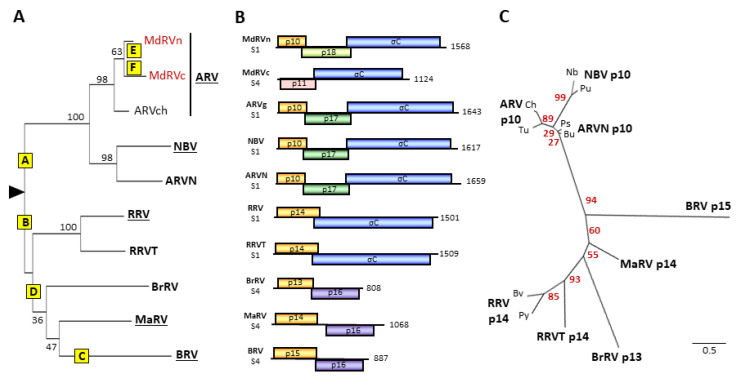
The family is the only non-enveloped virus family with members that use syncytium formation to promote cell-cell virus transmission. Syncytiogenesis is mediated by a fusion-associated small transmembrane (FAST) protein, a novel family of viral membrane fusion proteins. Previous evidence suggested the fusogenic reoviruses arose from an ancestral non-fusogenic virus, with the preponderance of fusogenic species suggesting positive evolutionary pressure to acquire and maintain the fusion phenotype. New phylogenetic analyses that included the atypical waterfowl subgroup of avian reoviruses and recently identified new orthoreovirus species indicate a more complex relationship between reovirus speciation and fusogenic capacity, with numerous predicted internal indels and 5'-terminal extensions driving the evolution of the orthoreovirus' polycistronic genome segments and their encoded FAST and fiber proteins. These inferred recombination events generated bi- and tricistronic genome segments with diverse gene constellations, they occurred pre- and post-orthoreovirus speciation, and they directly contributed to the evolution of the four extant orthoreovirus FAST proteins by driving both the gain and loss of fusion capability. We further show that two distinct post-speciation genetic events led to the loss of fusion in the waterfowl isolates of avian reovirus, a recombination event that replaced the p10 FAST protein with a heterologous, non-fusogenic protein and point substitutions in a conserved motif that destroyed the p10 assembly into multimeric fusion platforms.
该家族是唯一具有利用合胞体形成促进细胞间病毒传播成员的无包膜病毒家族。合胞体形成由融合相关的小跨膜(FAST)蛋白介导,这是一类新型的病毒膜融合蛋白。先前的证据表明,融合性呼肠孤病毒源自于祖先的非融合性病毒,大多数融合性物种表明,获得和维持融合表型存在正向进化压力。新的系统发育分析包括了禽呼肠孤病毒的非典型水禽亚群和最近鉴定的新正呼肠孤病毒种,这表明呼肠孤病毒的物种形成和融合能力之间存在更为复杂的关系,众多预测的内部缺失和 5'端延伸驱动了正呼肠孤病毒多顺反子基因组片段及其编码的 FAST 和纤维蛋白的进化。这些推断的重组事件产生了具有不同基因组合的双顺反子和三顺反子基因组片段,它们发生在正呼肠孤病毒物种形成之前和之后,并且通过驱动融合能力的获得和丧失,直接促进了四种现生正呼肠孤病毒 FAST 蛋白的进化。我们进一步表明,两个不同的种间遗传事件导致了水禽禽呼肠孤病毒分离株中融合能力的丧失,一个重组事件用异源的、非融合性蛋白取代了 p10 FAST 蛋白,另一个点突变破坏了 p10 组装成多聚体融合平台的保守模体。