Howard Hughes Medical Institute, Institute of Neuroscience, University of Oregon, Eugene, OR, 97403, USA.
Neural Dev. 2020 Jul 28;15(1):9. doi: 10.1186/s13064-020-00146-6.
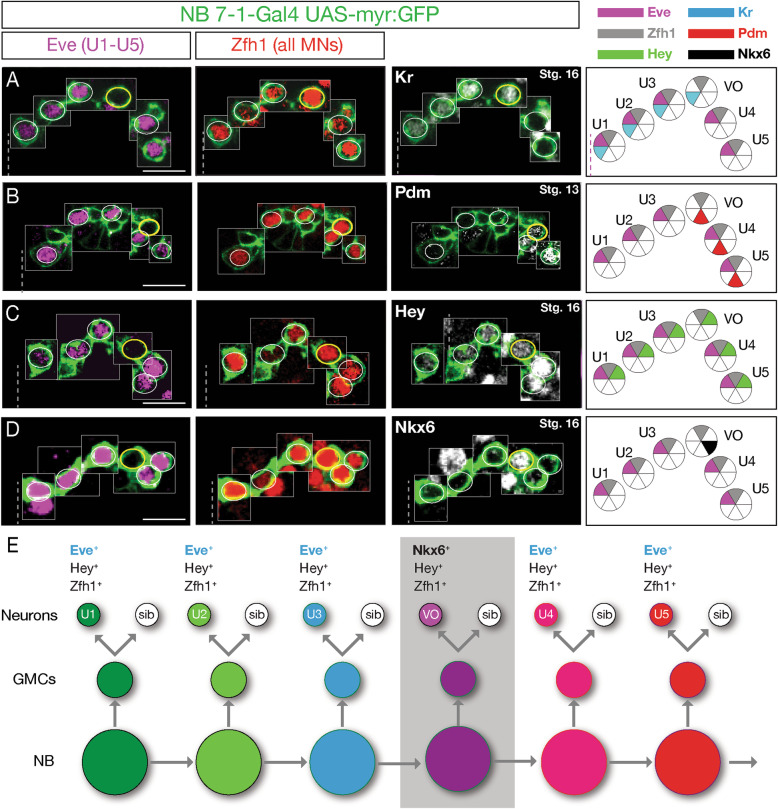
Spatial patterning specifies neural progenitor identity, with further diversity generated by temporal patterning within individual progenitor lineages. In vertebrates, these mechanisms generate "cardinal classes" of neurons that share a transcription factor identity and common morphology. In Drosophila, two cardinal classes are Even-skipped (Eve) motor neurons projecting to dorsal longitudinal muscles, and Nkx6 motor neurons projecting to ventral oblique muscles. Cross-repressive interactions prevent stable double-positive motor neurons. The Drosophila neuroblast 7-1 (NB7-1) lineage uses a temporal transcription factor cascade to generate five distinct Eve motor neurons; the origin and development of Nkx6 motor neurons remains unclear.
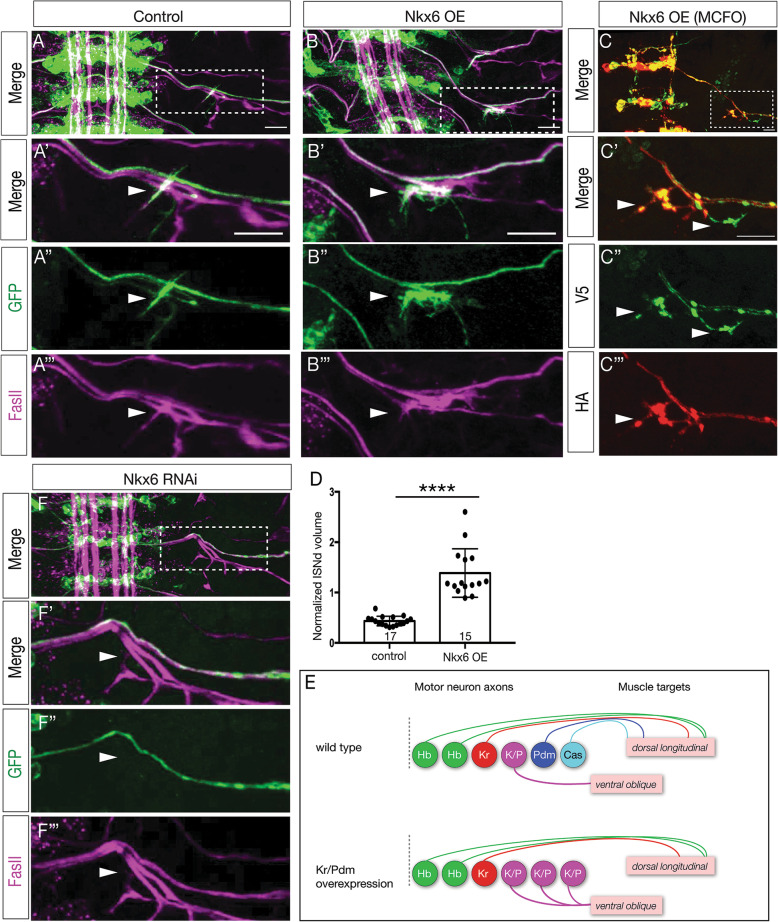
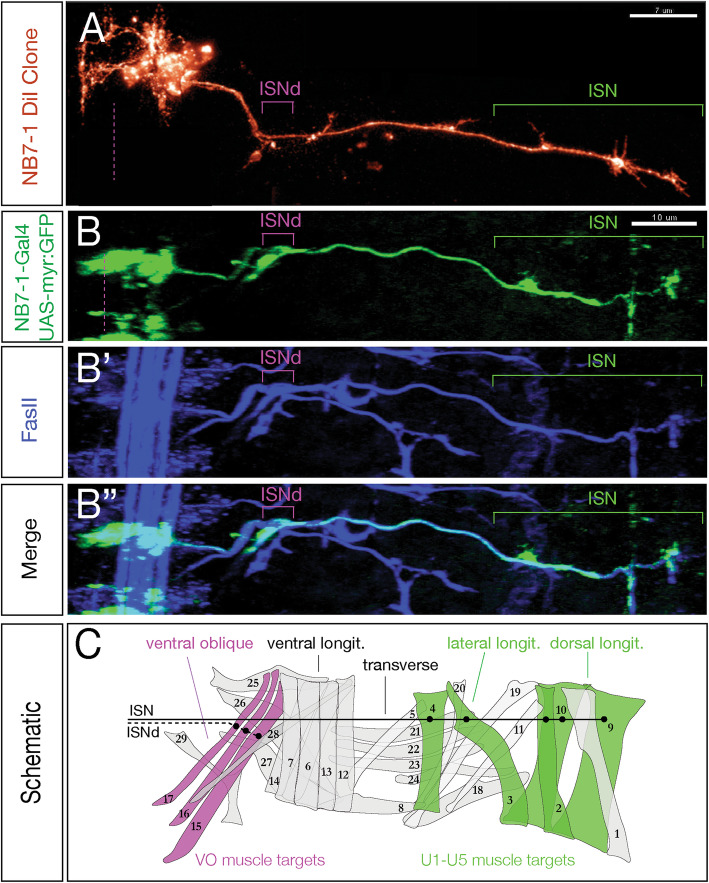
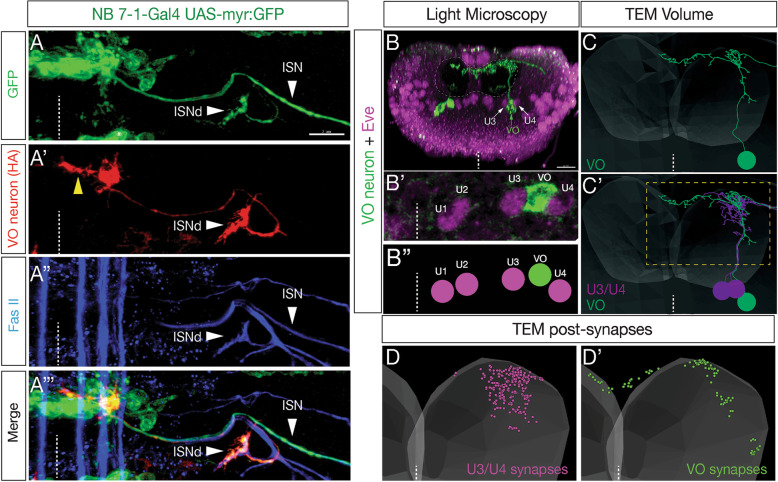
We use a neuroblast specific Gal4 line, sparse labelling and molecular markers to identify an Nkx6 VO motor neuron produced by the NB7-1 lineage. We use lineage analysis to birth-date the VO motor neuron to the Kr Pdm neuroblast temporal identity window. We use gain- and loss-of-function strategies to test the role of Kr Pdm temporal identity and the Nkx6 transcription factor in specifying VO neuron identity.
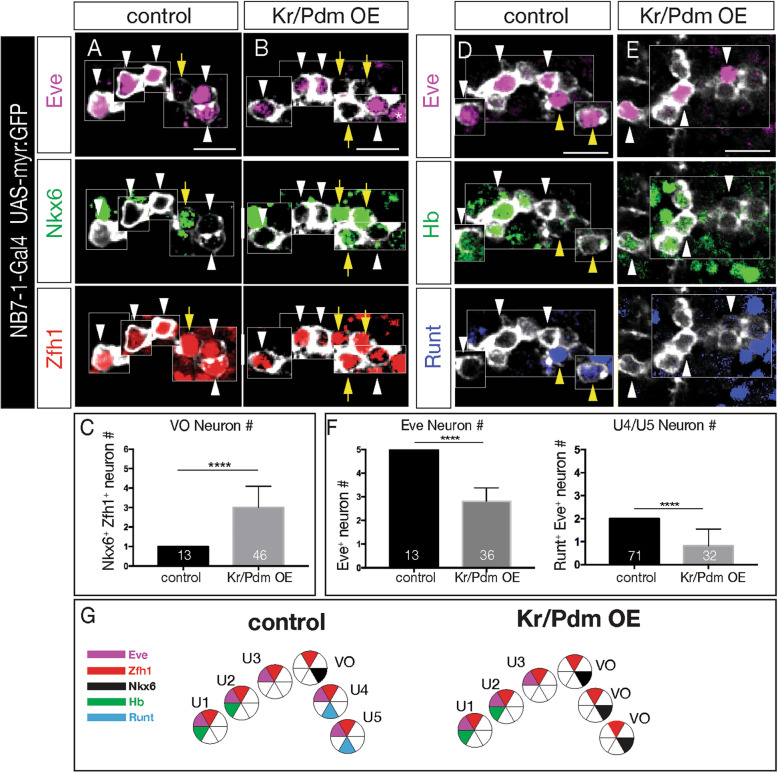
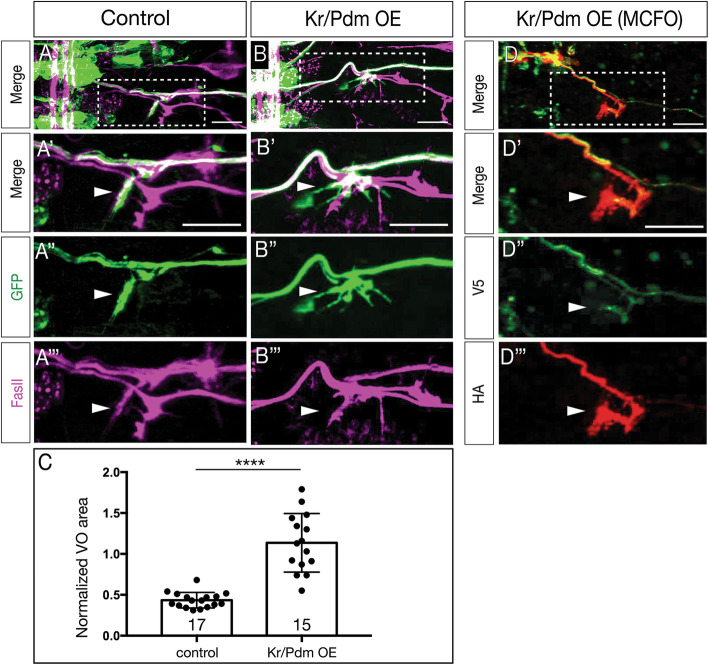
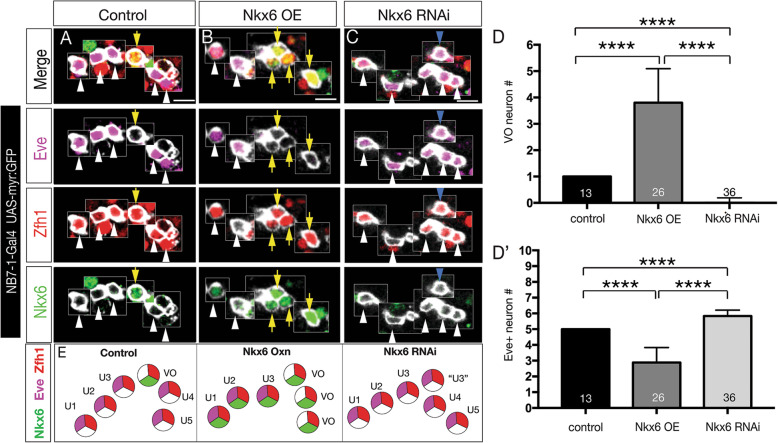
Lineage analysis identifies an Nkx6 neuron born from the Kr Pdm temporal identity window in the NB7-1 lineage, resulting in alternation of cardinal motor neuron subtypes within this lineage (Eve>Nkx6 > Eve). Co-overexpression of Kr/Pdm generates ectopic VO motor neurons within the NB7-1 lineage - the first evidence that this TTF combination specifies neuronal identity. Moreover, the Kr/Pdm combination promotes Nkx6 expression, which itself is necessary and sufficient for motor neuron targeting to ventral oblique muscles, thereby revealing a molecular specification pathway from temporal patterning to cardinal transcription factor expression to motor neuron target selection.
We show that one neuroblast lineage generates interleaved cardinal motor neurons fates; that the Kr/Pdm TTFs form a novel temporal identity window that promotes expression of Nkx6; and that the Kr/Pdm > Nkx6 pathway is necessary and sufficient to promote VO motor neuron targeting to the correct ventral muscle group.
空间模式决定神经祖细胞的身份,而个体祖细胞谱系内的时间模式进一步产生多样性。在脊椎动物中,这些机制产生了具有转录因子身份和共同形态的“主要神经元类群”。在果蝇中,两类主要神经元是投射到背纵肌的 Even-skipped(Eve)运动神经元,以及投射到腹斜肌的 Nkx6 运动神经元。交叉抑制相互作用阻止了稳定的双阳性运动神经元的产生。果蝇神经母细胞 7-1(NB7-1)谱系使用时间转录因子级联产生五个不同的 Eve 运动神经元;Nkx6 运动神经元的起源和发育仍不清楚。
我们使用一种神经母细胞特异性 Gal4 系,稀疏标记和分子标记来鉴定由 NB7-1 谱系产生的 Nkx6 VO 运动神经元。我们使用谱系分析来确定 VO 运动神经元的出生日期到 Kr Pdm 神经母细胞的时间身份窗口。我们使用增益和缺失功能策略来测试 Kr Pdm 时间身份和 Nkx6 转录因子在指定 VO 神经元身份中的作用。
谱系分析确定了由 NB7-1 谱系中的 Kr Pdm 时间身份窗口产生的 Nkx6 神经元,导致该谱系内主要运动神经元亚型的交替(Eve>Nkx6>Eve)。Kr/Pdm 的共过表达在 NB7-1 谱系内产生了异位 VO 运动神经元——这是第一个证明这种 TTF 组合指定神经元身份的证据。此外,Kr/Pdm 组合促进了 Nkx6 的表达,这本身对于运动神经元靶向腹斜肌是必要和充分的,从而揭示了从时间模式到主要转录因子表达再到运动神经元靶选择的分子指定途径。
我们表明,一个神经母细胞谱系产生了交错的主要运动神经元命运;Kr/Pdm TTFs 形成了一个新的时间身份窗口,促进了 Nkx6 的表达;并且 Kr/Pdm>Nkx6 途径是促进 VO 运动神经元靶向正确的腹肌群的必要和充分条件。