ISGlobal, Hospital Clinic - Universitat de Barcelona, Barcelona, Spain.
Department of Biomedical Sciences, Institute of Tropical Medicine, Antwerp, Belgium.
Elife. 2020 Oct 21;9:e60058. doi: 10.7554/eLife.60058.
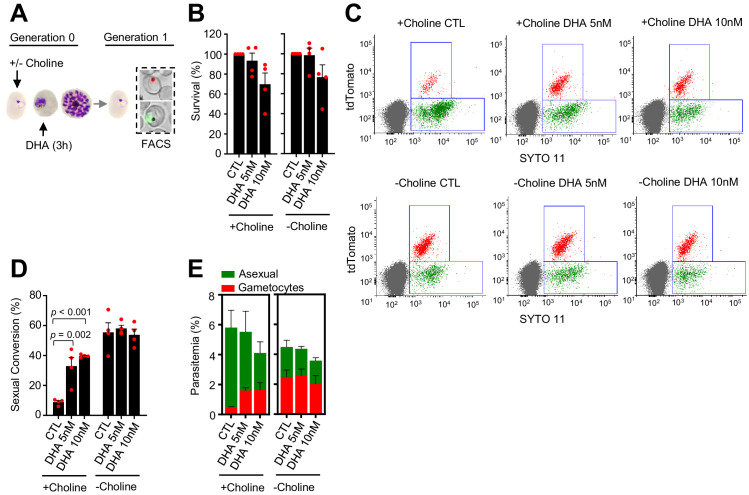
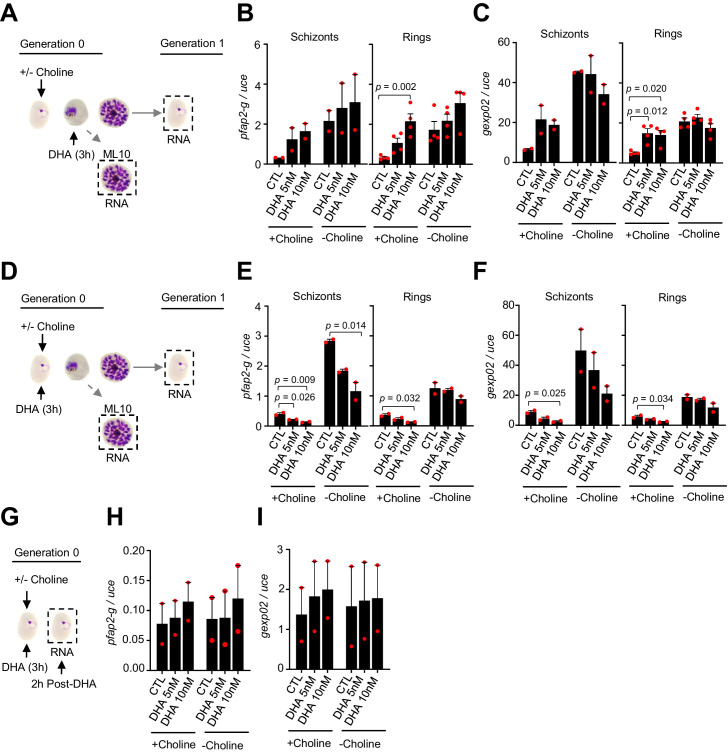
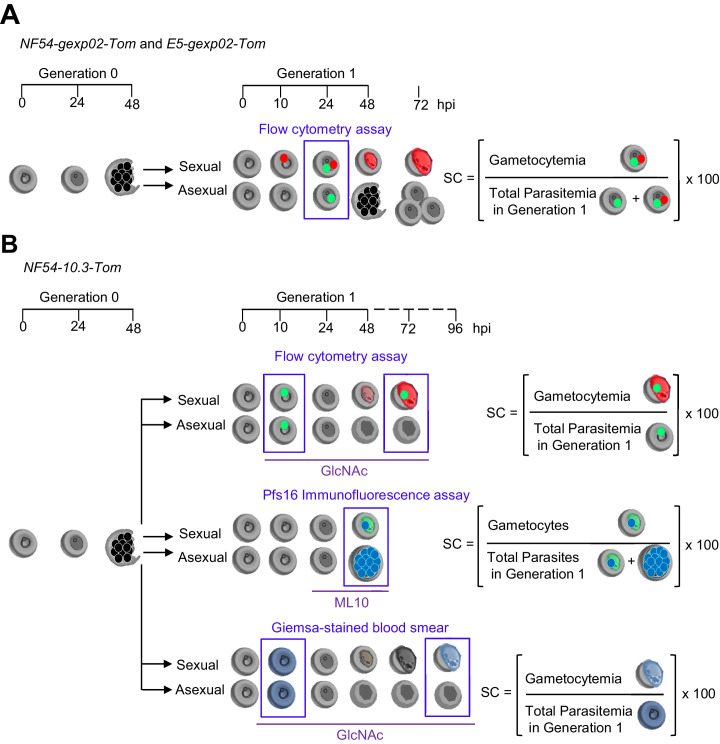
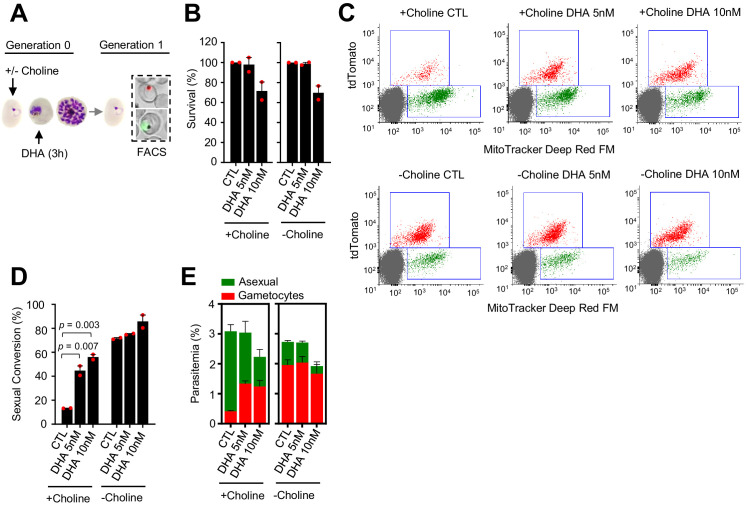
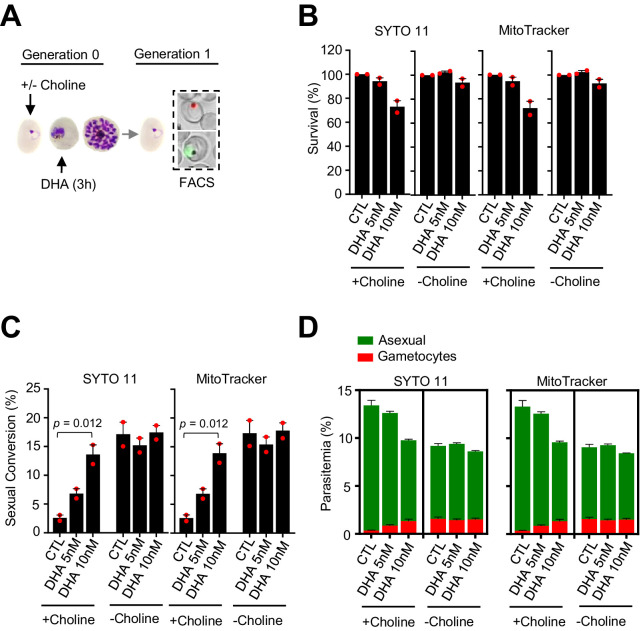
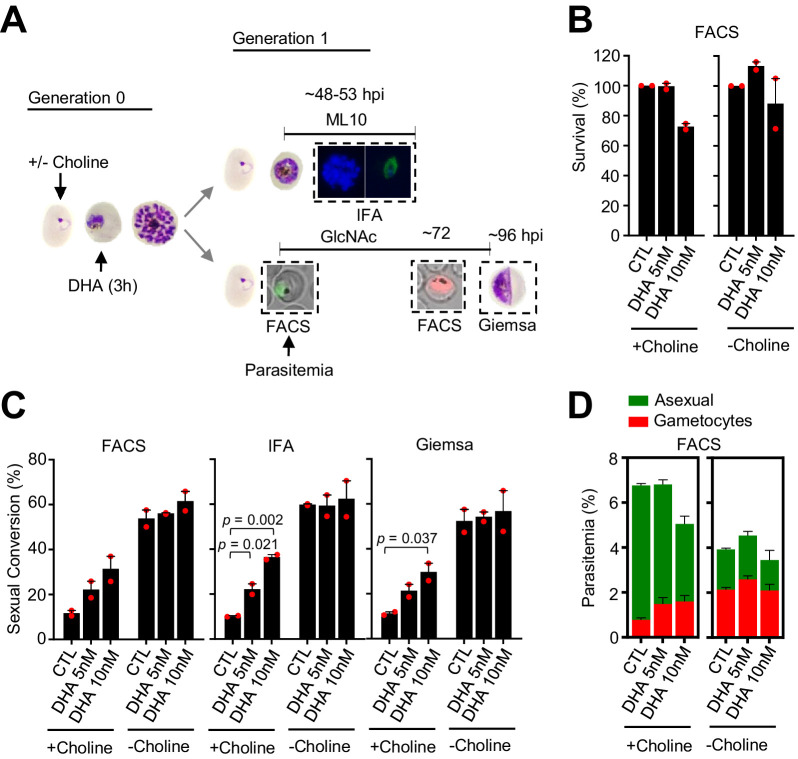
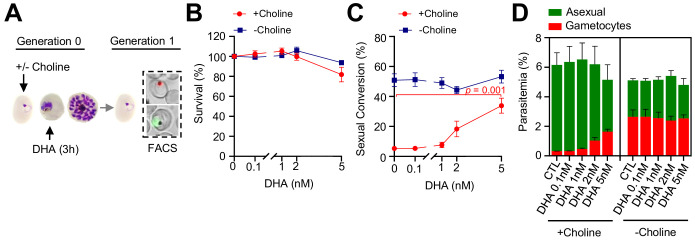
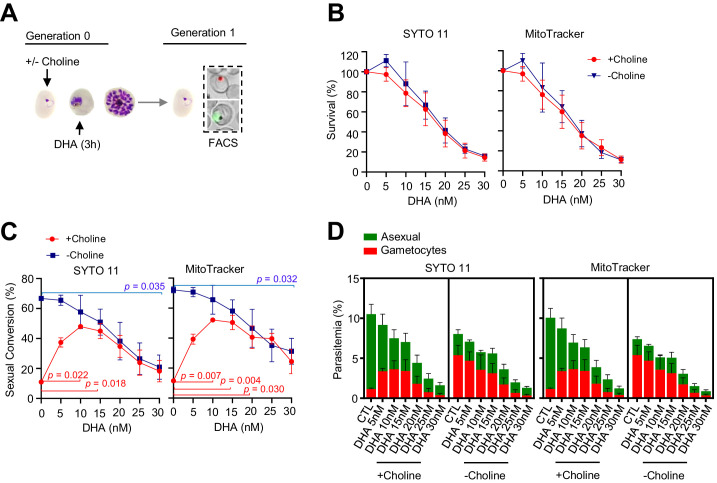
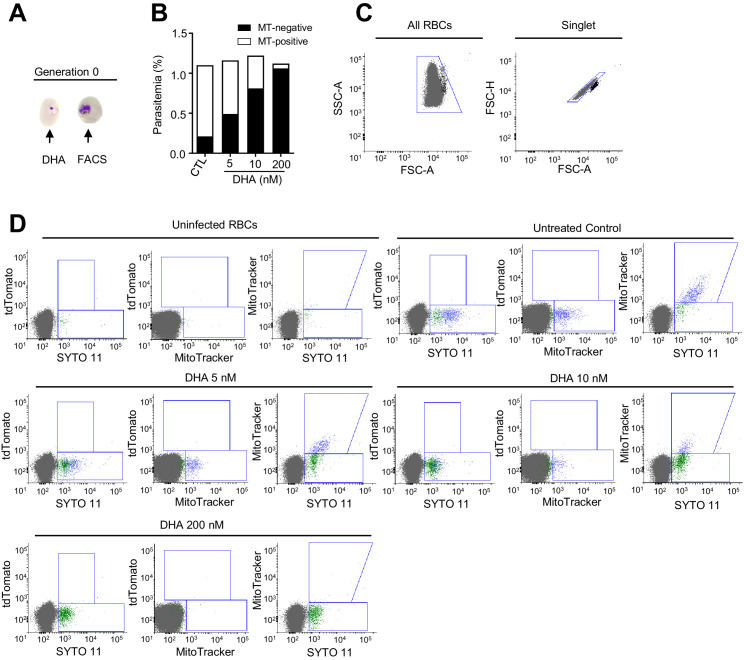
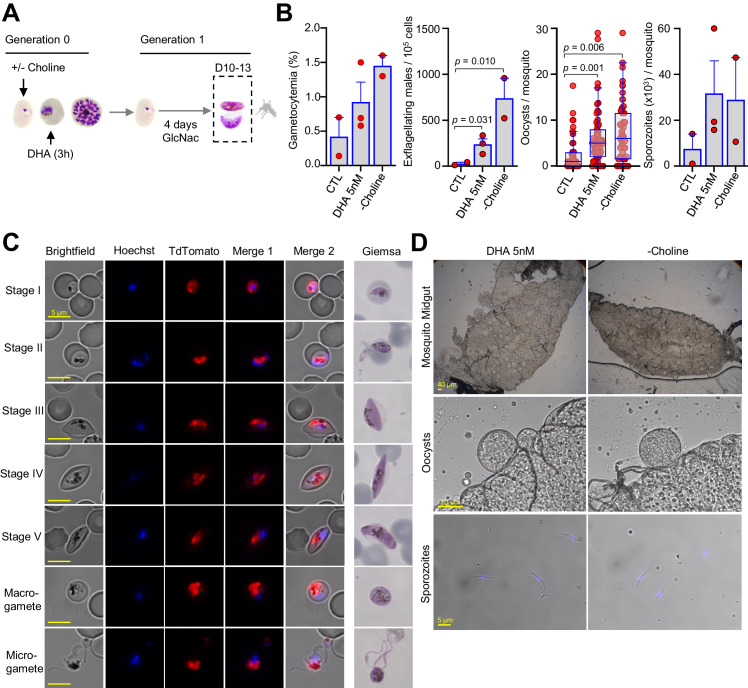
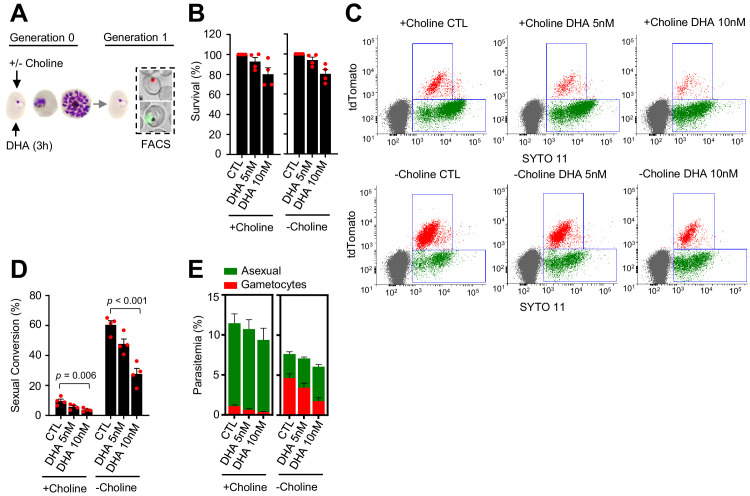
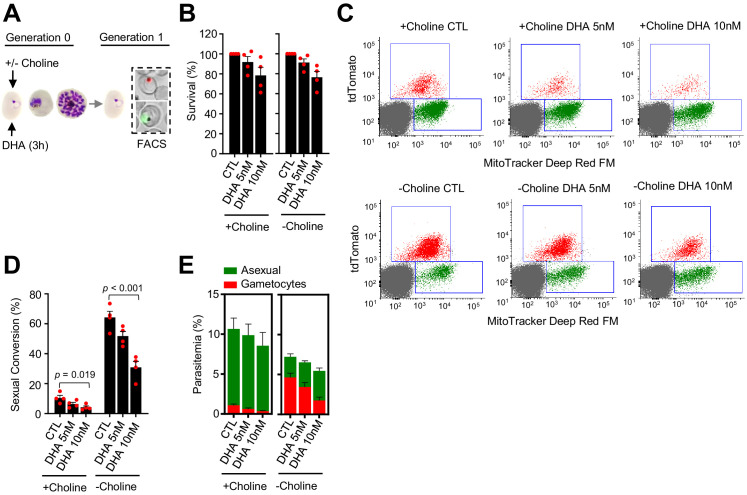
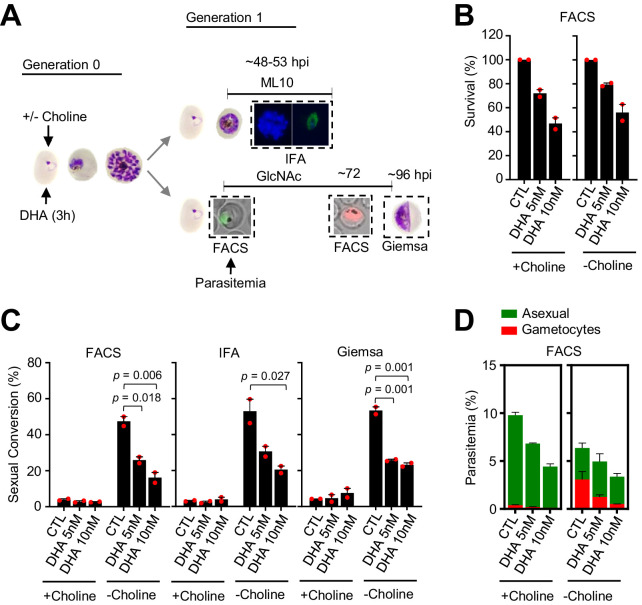
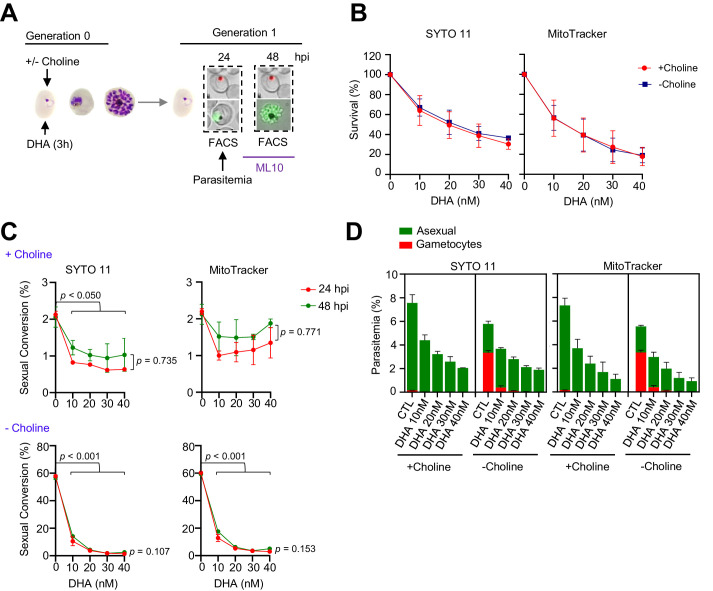
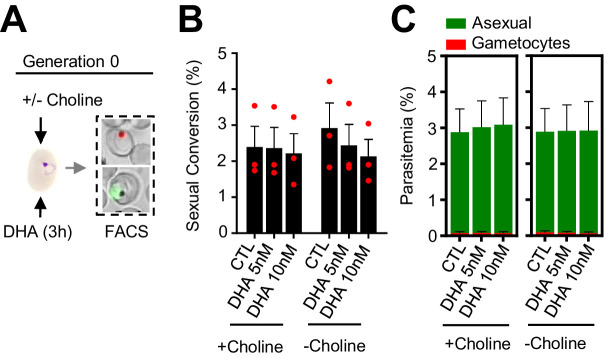
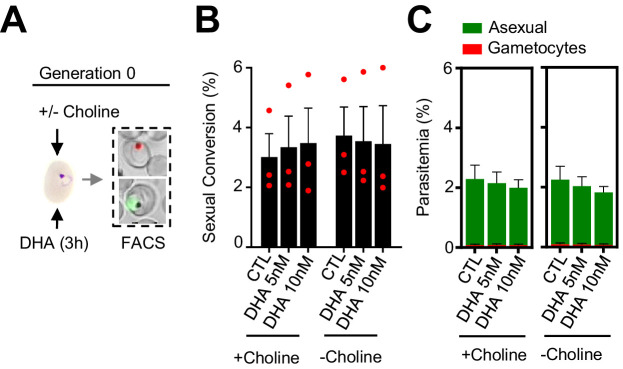
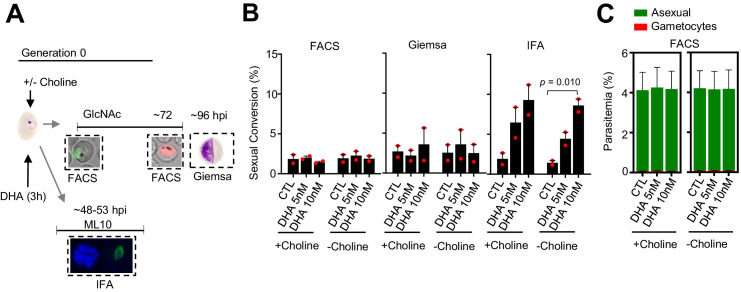
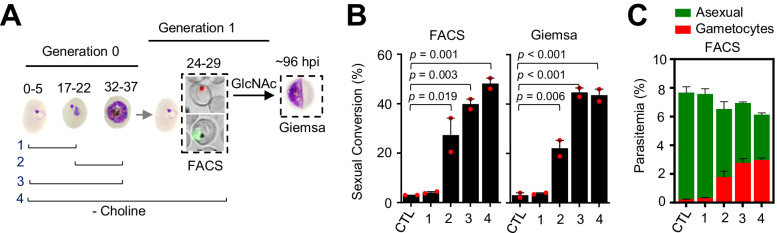
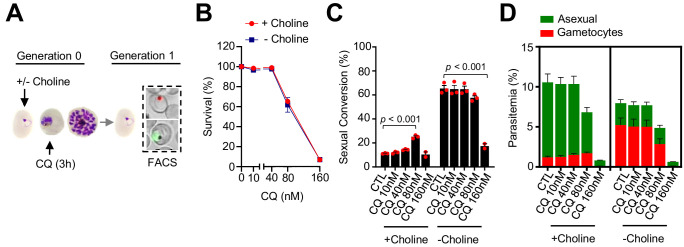
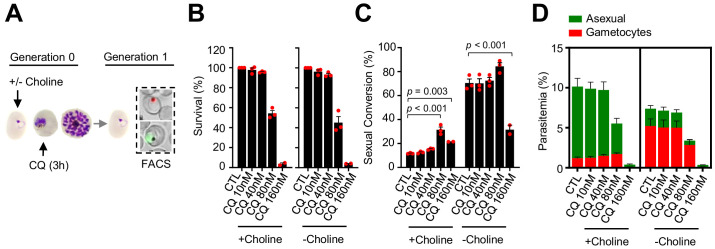
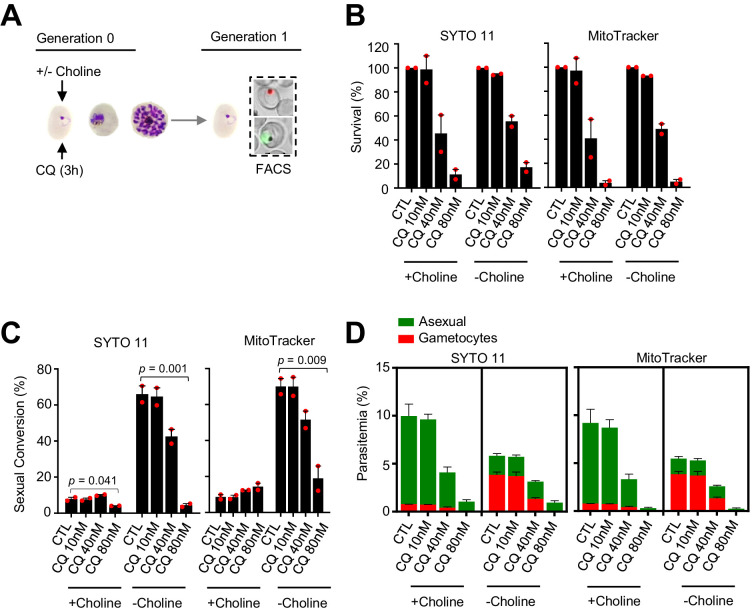
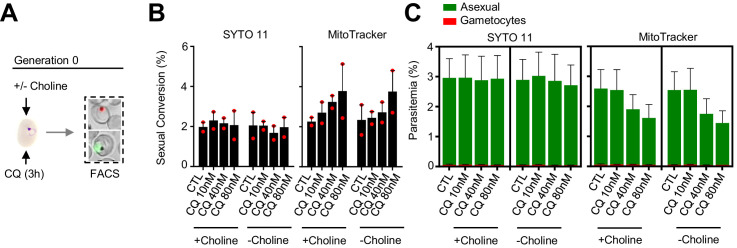
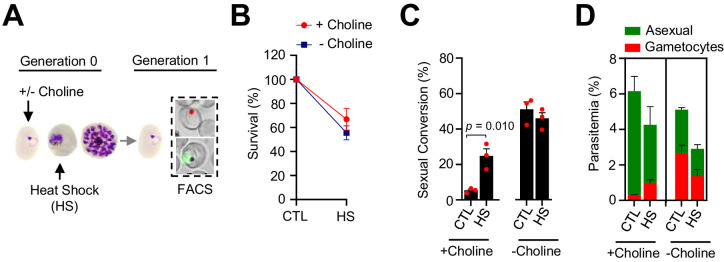
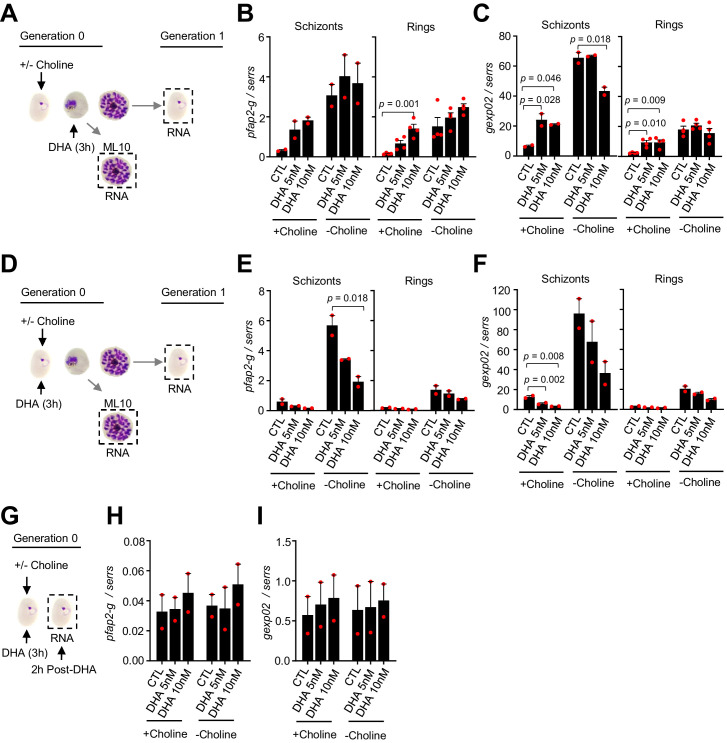
Malaria transmission is dependent on the formation of gametocytes in the human blood. The sexual conversion rate, the proportion of asexual parasites that convert into gametocytes at each multiplication cycle, is variable and reflects the relative parasite investment between transmission and maintaining the infection. The impact of environmental factors such as drugs on sexual conversion rates is not well understood. We developed a robust assay using gametocyte-reporter parasite lines to accurately measure the impact of drugs on sexual conversion rates, independently from their gametocytocidal activity. We found that exposure to subcurative doses of the frontline antimalarial drug dihydroartemisinin (DHA) at the trophozoite stage resulted in a ~ fourfold increase in sexual conversion. In contrast, no increase was observed when ring stages were exposed or in cultures in which sexual conversion was stimulated by choline depletion. Our results reveal a complex relationship between antimalarial drugs and sexual conversion, with potential public health implications.
疟疾传播依赖于人体血液中配子体的形成。性转化率,即每个增殖周期中无性寄生虫转化为配子体的比例是可变的,反映了寄生虫在传播和维持感染之间的相对投入。环境因素(如药物)对性转化率的影响尚不清楚。我们开发了一种使用配子体报告寄生虫系的稳健测定法,可准确测量药物对性转化率的影响,而与它们的配子体杀伤活性无关。我们发现,在前驱抗疟药物二氢青蒿素(DHA)的滋养体阶段暴露于亚治疗剂量会导致性转化增加约四倍。相比之下,当环阶段暴露或在通过胆碱耗竭刺激性转化的培养物中,未观察到增加。我们的结果揭示了抗疟药物与性转化之间的复杂关系,这可能对公共卫生具有重要意义。