Department of Medicine, University of Chicago, Chicago, United States.
Department of Human Genetics, University of Chicago, Chicago, United States.
Elife. 2020 Oct 21;9:e59929. doi: 10.7554/eLife.59929.
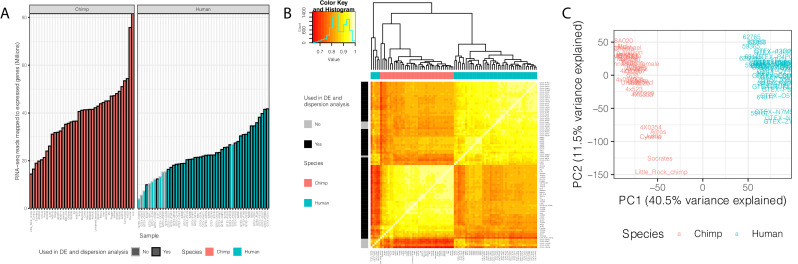
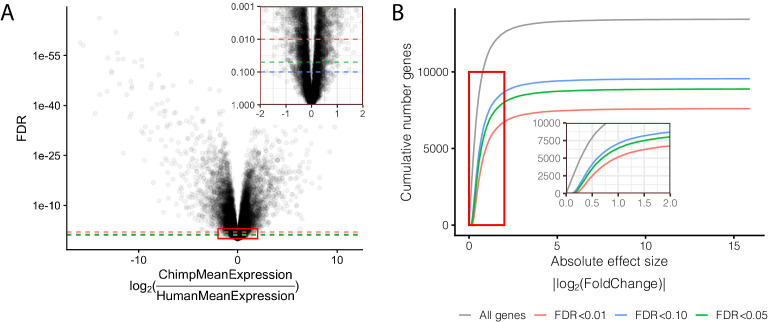
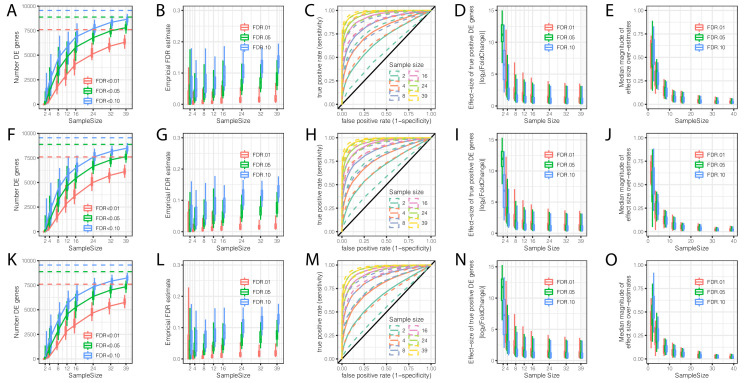
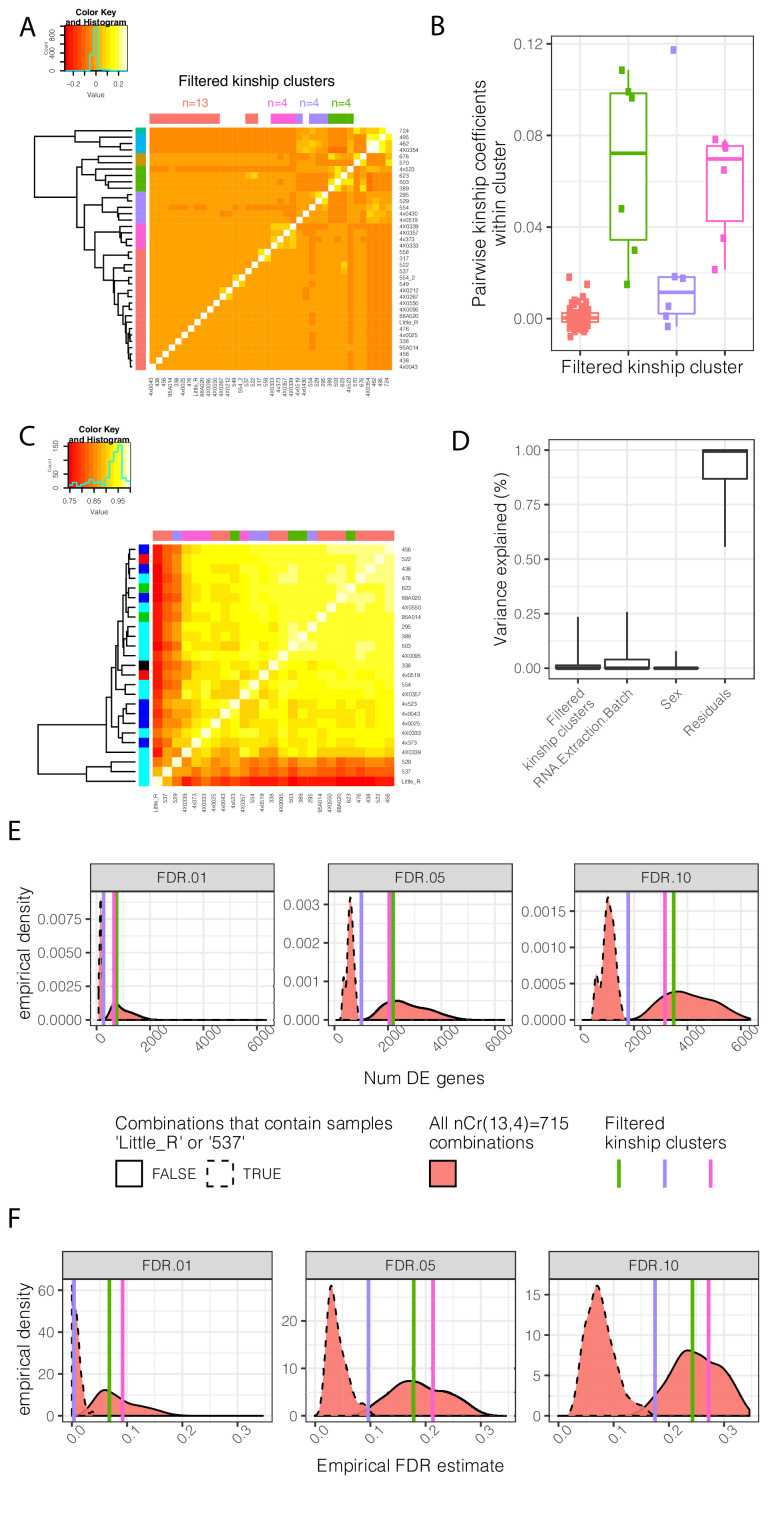
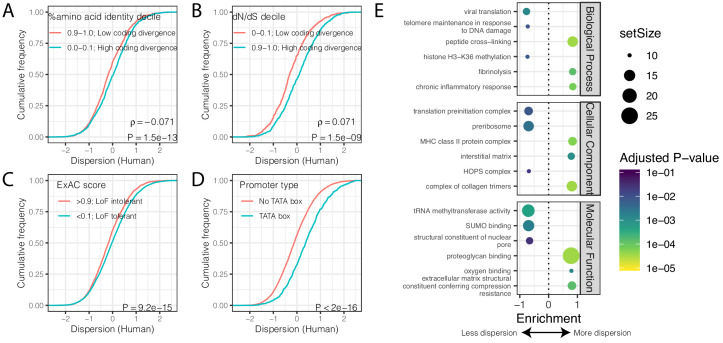
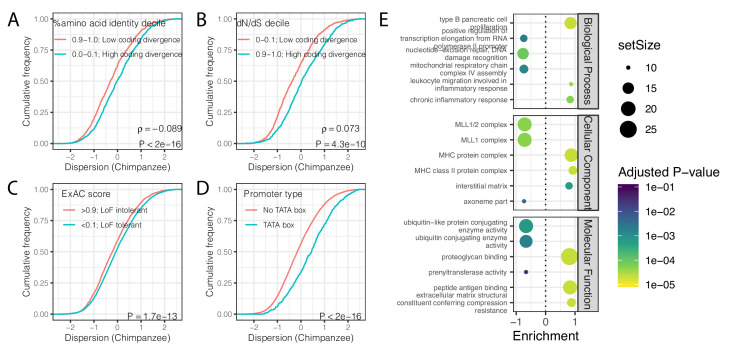
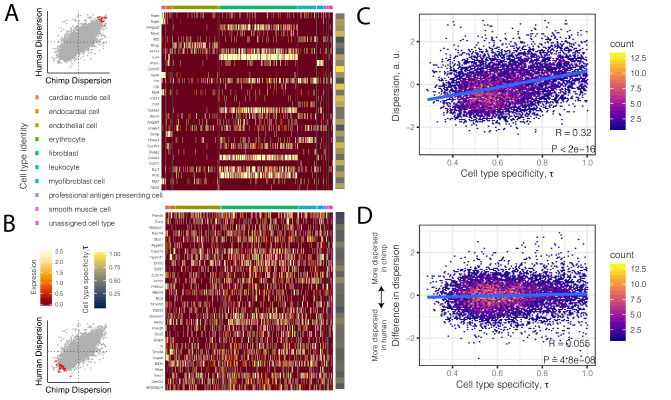
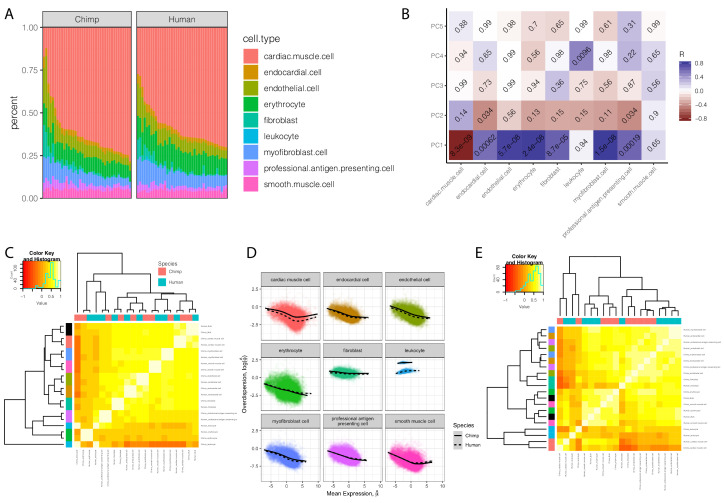
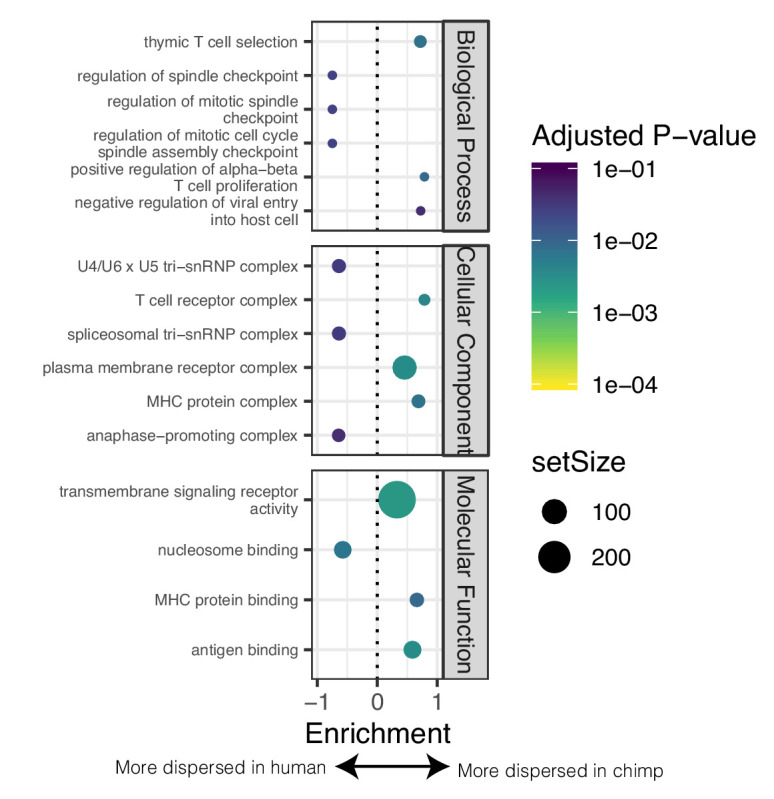
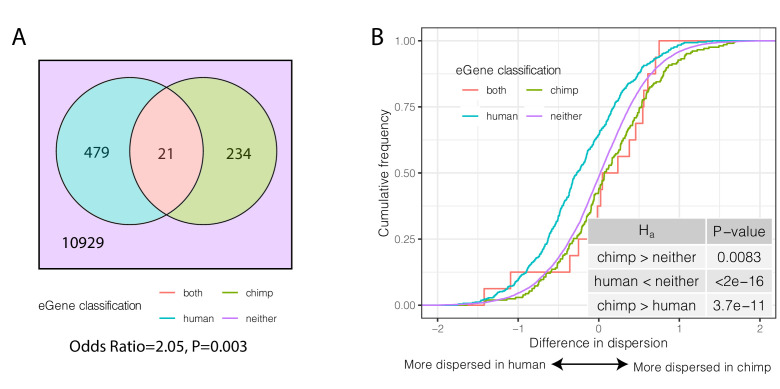
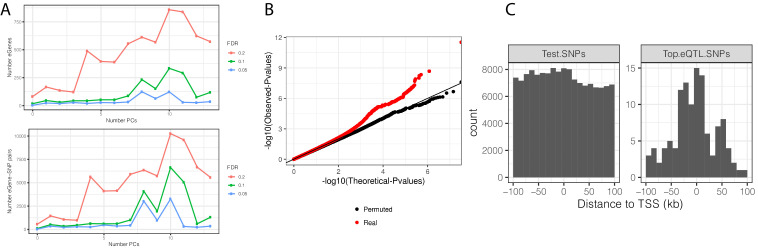
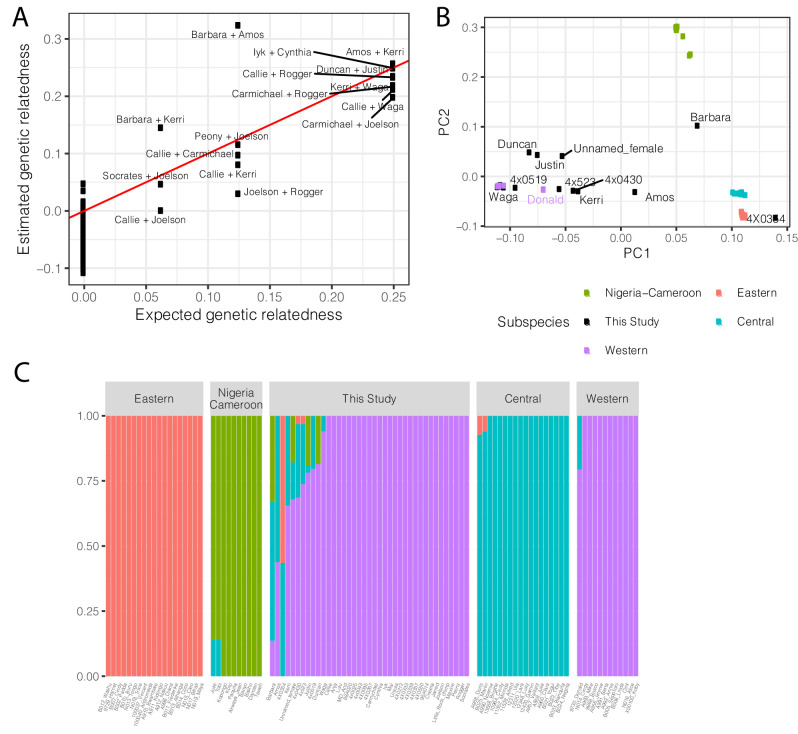
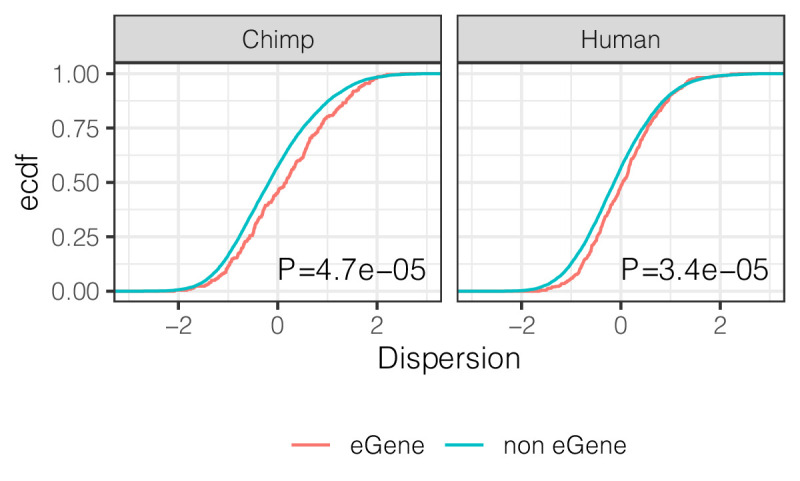
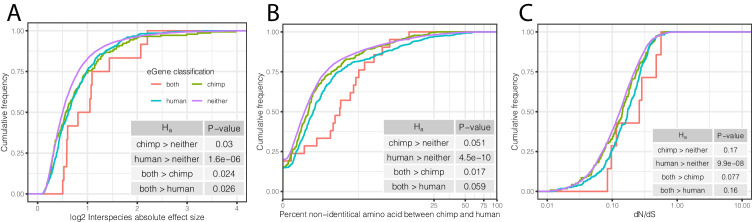
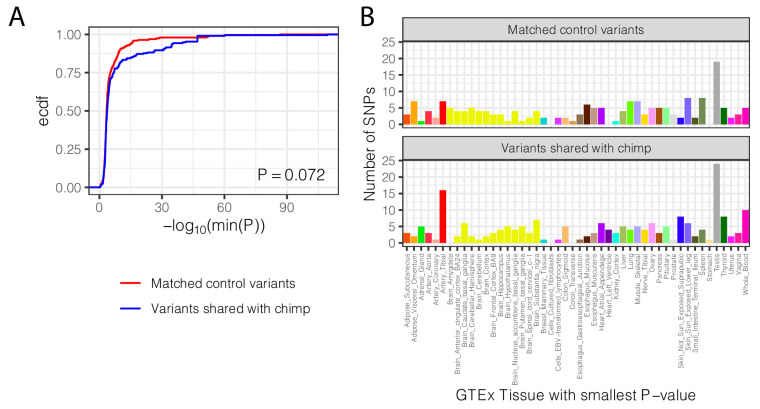
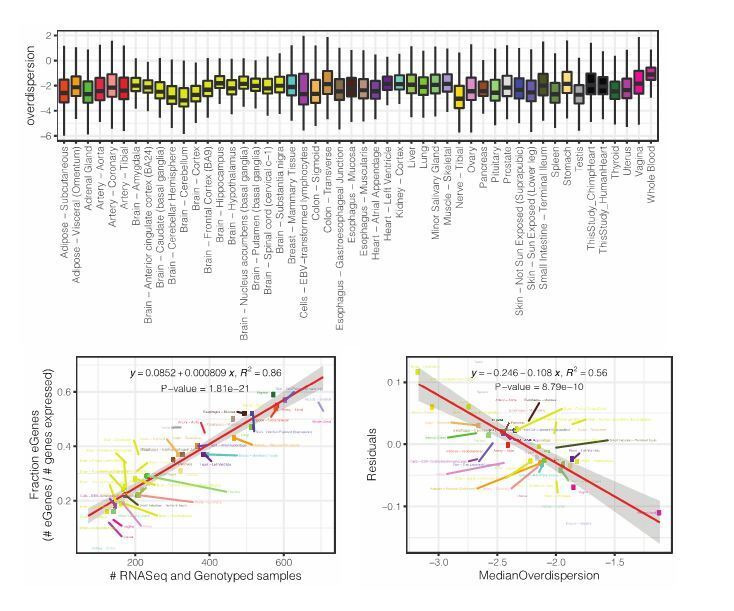
Inter-individual variation in gene expression has been shown to be heritable and is often associated with differences in disease susceptibility between individuals. Many studies focused on mapping associations between genetic and gene regulatory variation, yet much less attention has been paid to the evolutionary processes that shape the observed differences in gene regulation between individuals in humans or any other primate. To begin addressing this gap, we performed a comparative analysis of gene expression variability and expression quantitative trait loci (eQTLs) in humans and chimpanzees, using gene expression data from primary heart samples. We found that expression variability in both species is often determined by non-genetic sources, such as cell-type heterogeneity. However, we also provide evidence that inter-individual variation in gene regulation can be genetically controlled, and that the degree of such variability is generally conserved in humans and chimpanzees. In particular, we found a significant overlap of orthologous genes associated with eQTLs in both species. We conclude that gene expression variability in humans and chimpanzees often evolves under similar evolutionary pressures.
个体间基因表达的变化是可遗传的,通常与个体间疾病易感性的差异有关。许多研究集中于绘制遗传和基因调控变异之间的关联,但对于塑造人类或任何其他灵长类动物个体间基因调控差异的进化过程,关注较少。为了开始解决这一差距,我们使用来自原代心脏样本的基因表达数据,在人类和黑猩猩中进行了基因表达可变性和表达数量性状基因座(eQTLs)的比较分析。我们发现,两种物种的表达可变性通常由非遗传因素决定,如细胞类型异质性。然而,我们也提供了证据表明,基因调控的个体间差异可以受到遗传控制,并且这种可变性的程度在人类和黑猩猩中通常是保守的。特别是,我们发现了与两个物种中 eQTL 相关的同源基因的显著重叠。我们得出结论,人类和黑猩猩的基因表达可变性通常在相似的进化压力下进化。