Division of Cellular Medicine, School of Medicine, University of Dundee, Dundee, UK.
J Cell Biol. 2020 Dec 7;219(12). doi: 10.1083/jcb.202002020.
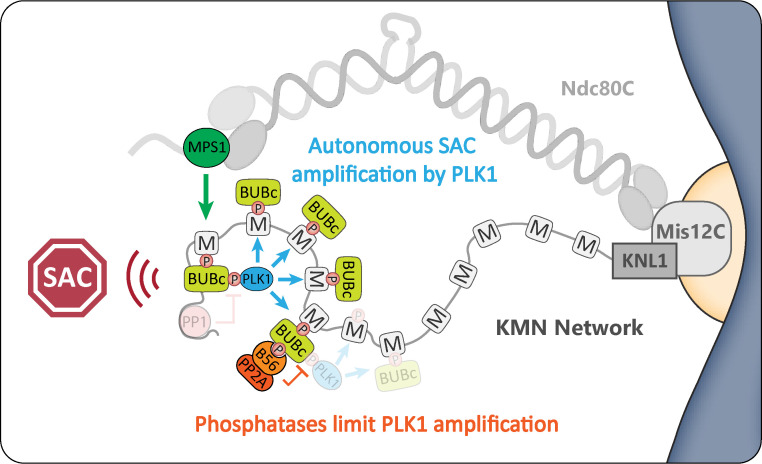
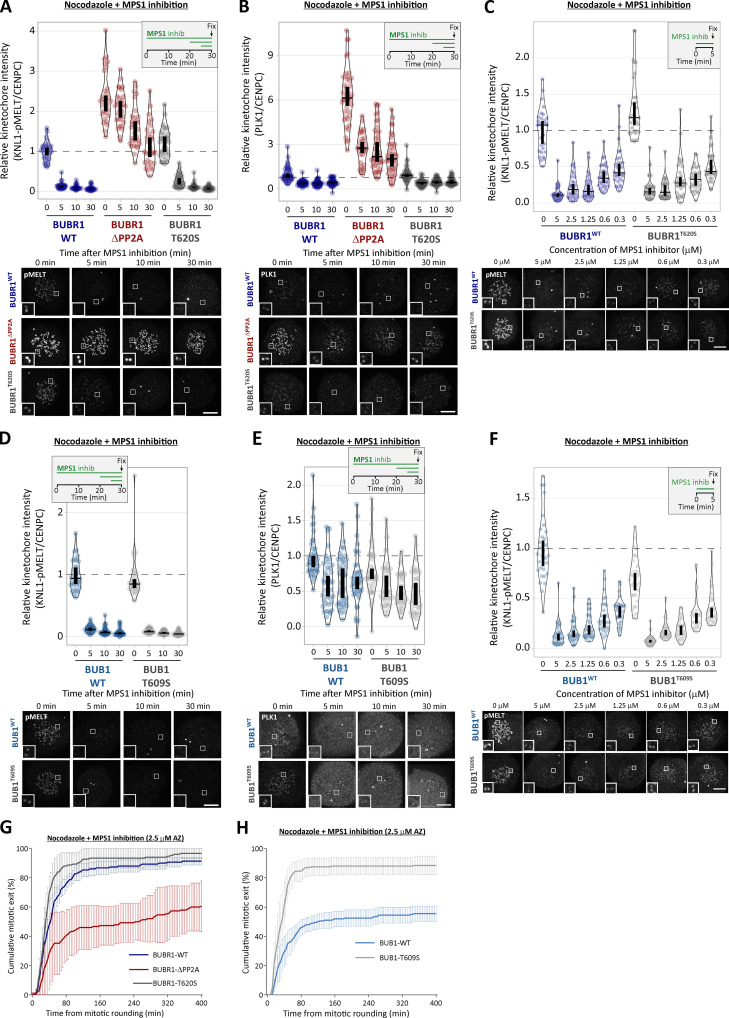
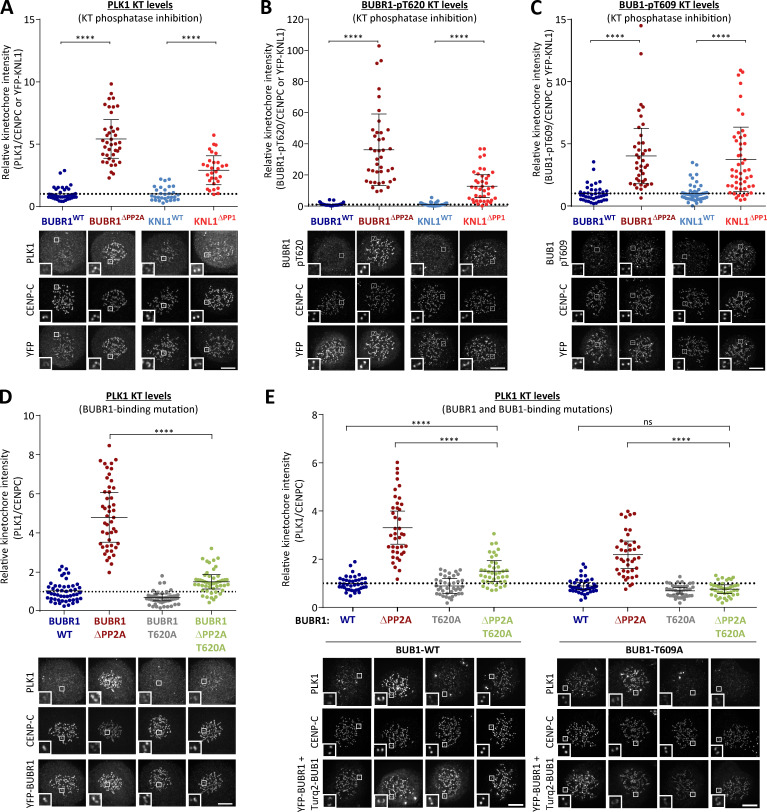
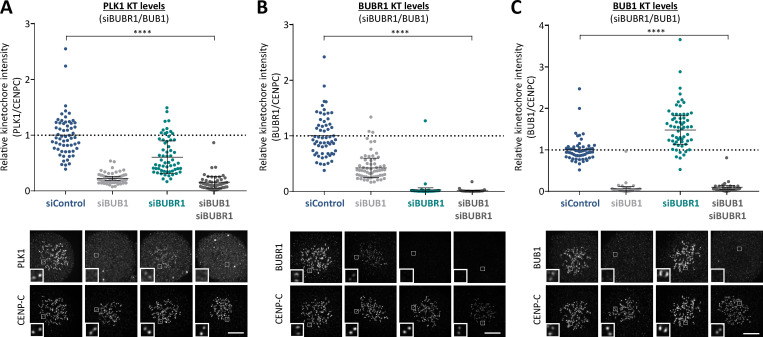
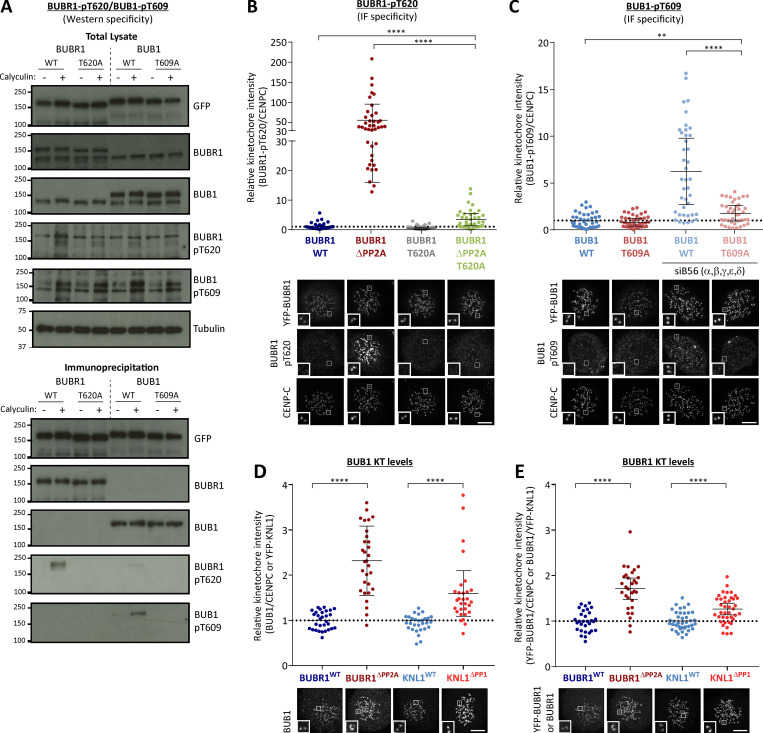
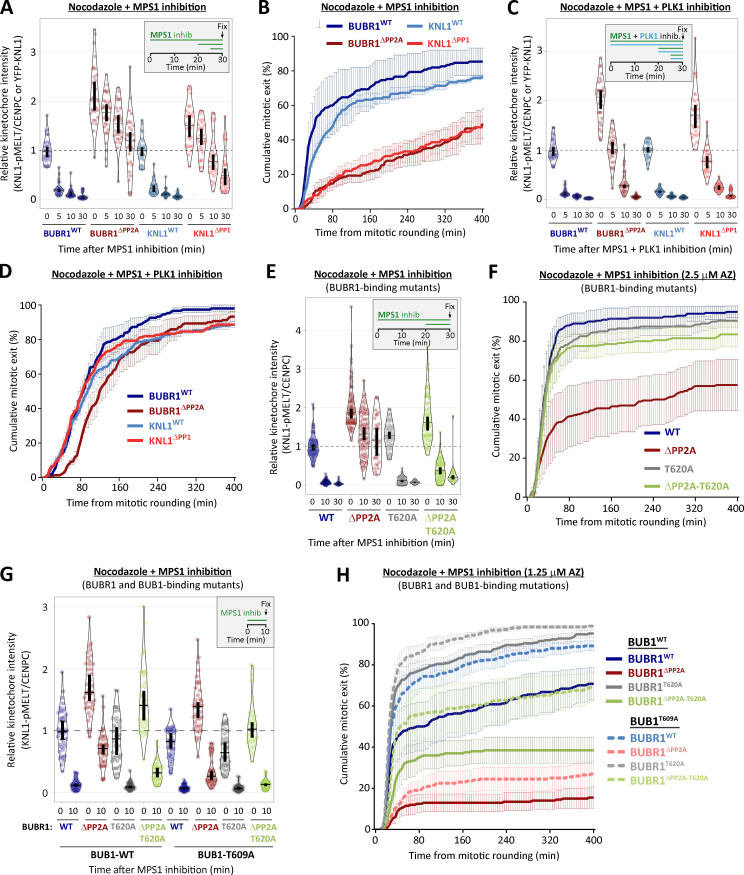
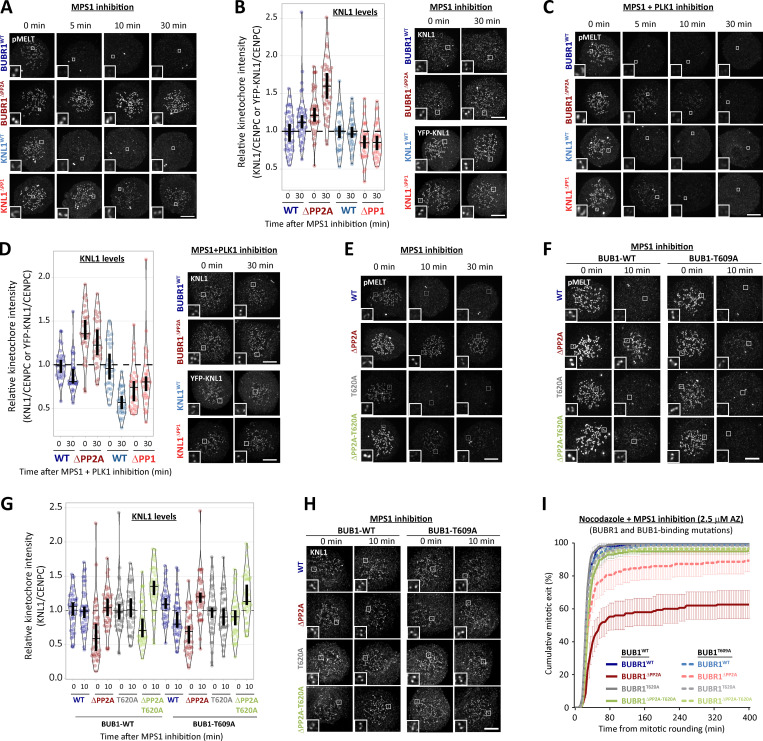
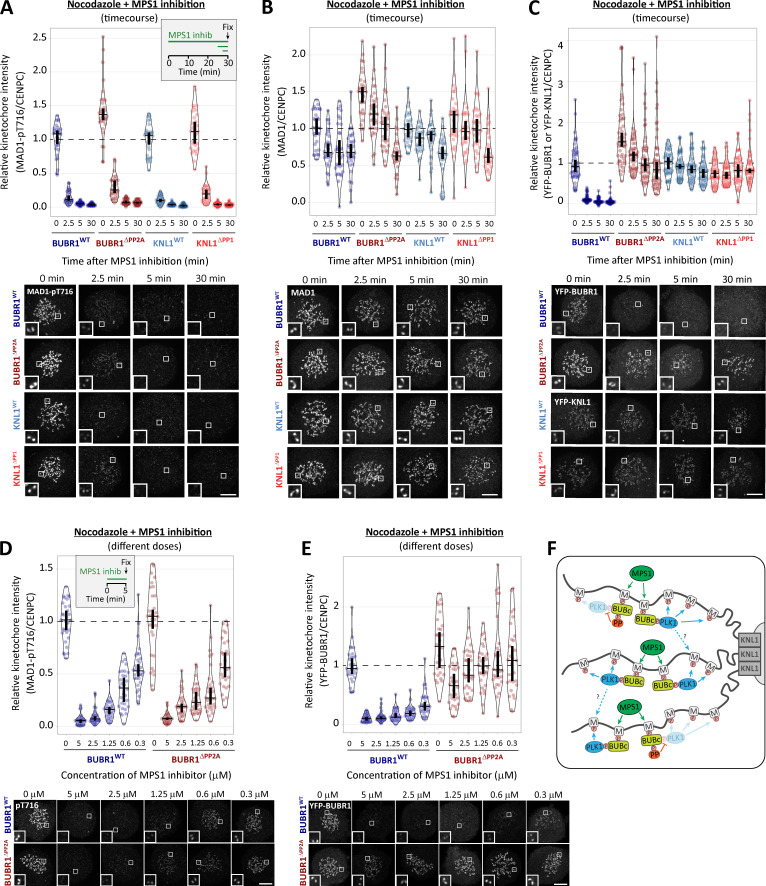
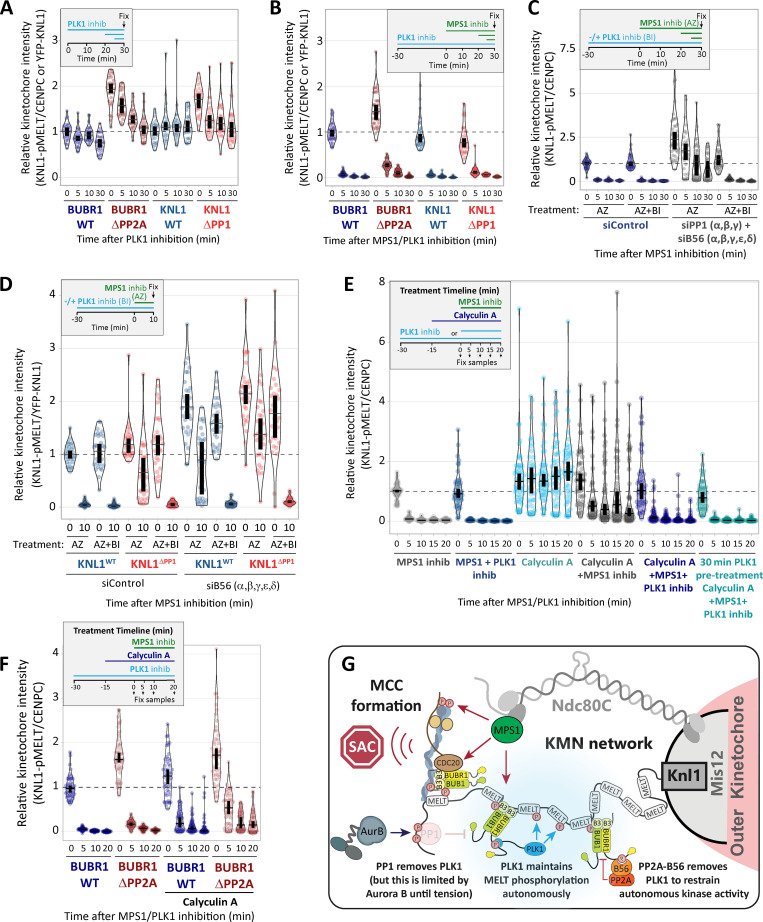
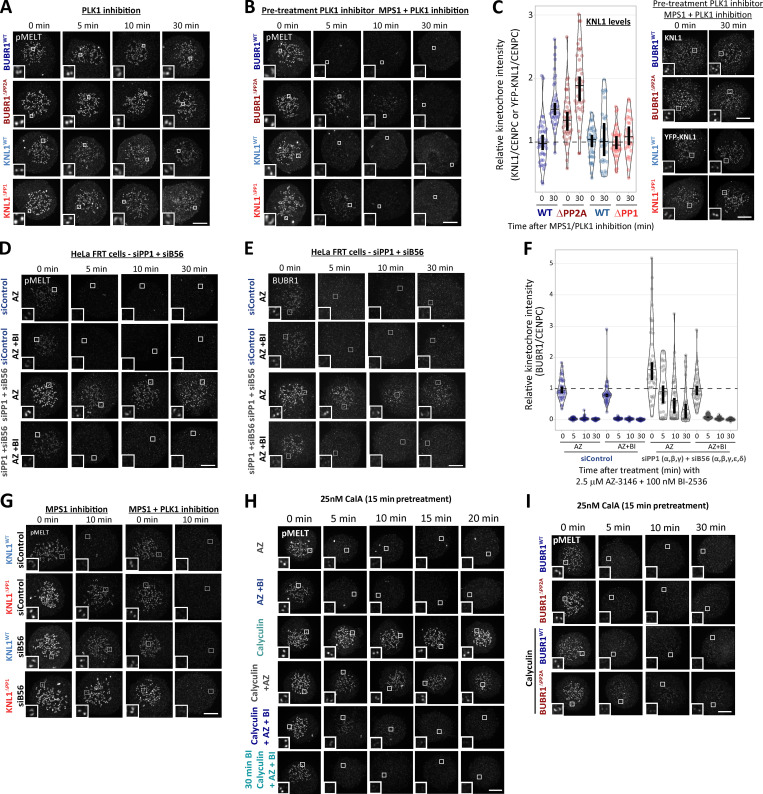
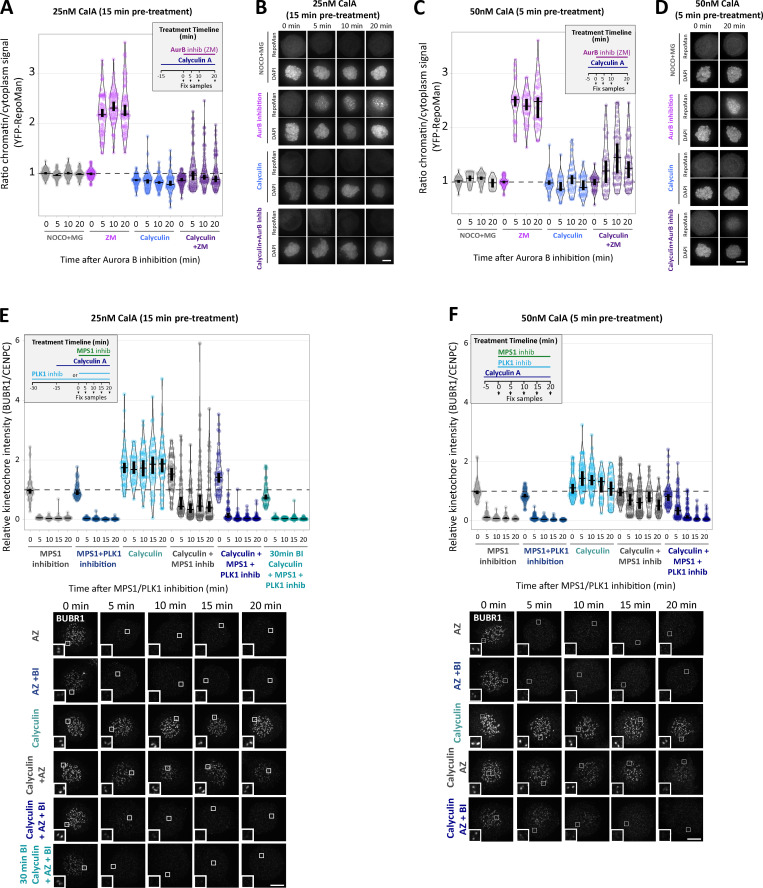
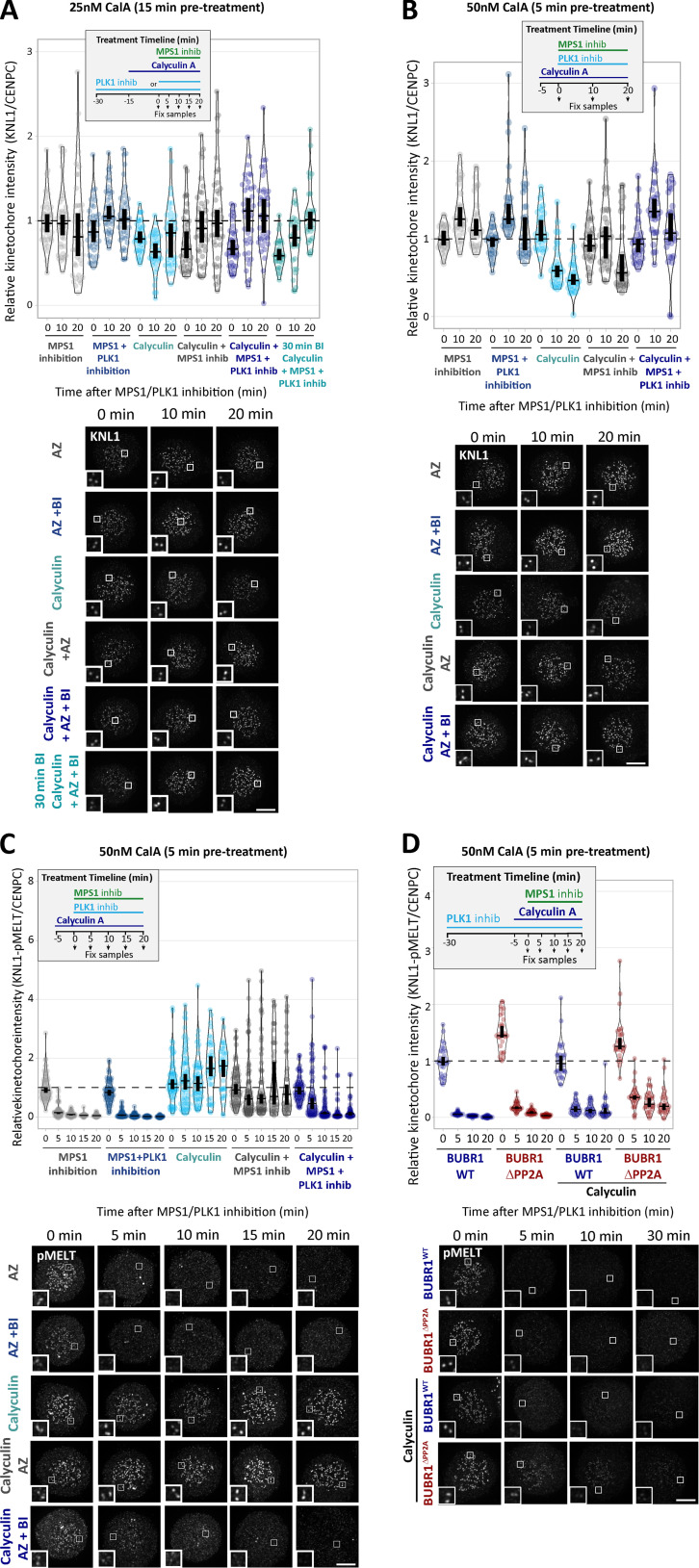
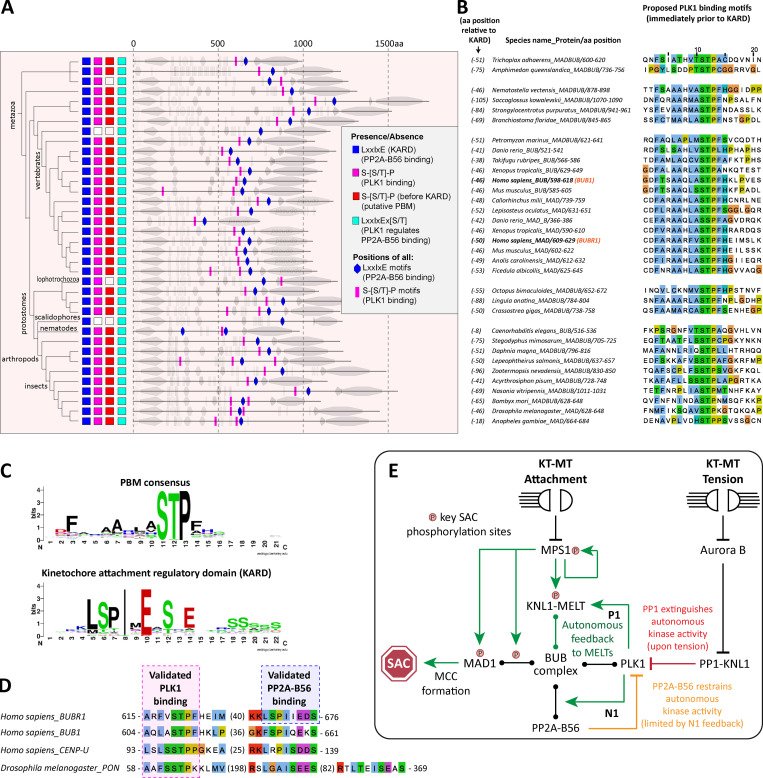
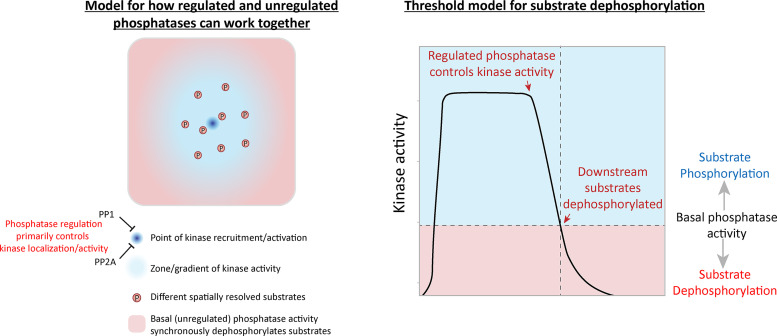
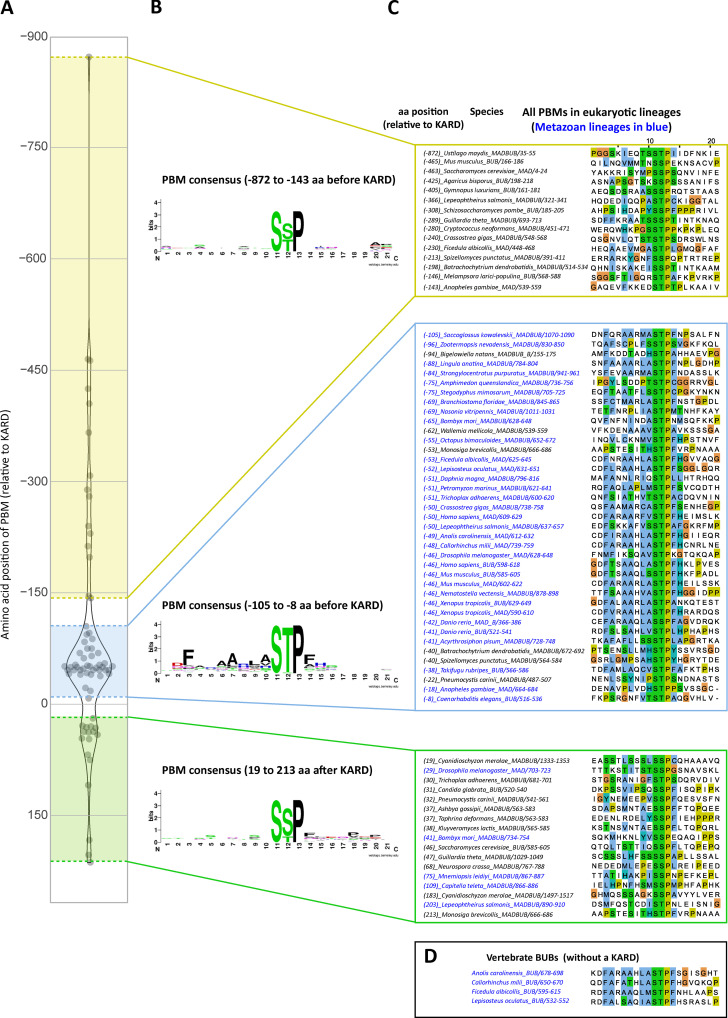
Local phosphatase regulation is needed at kinetochores to silence the mitotic checkpoint (a.k.a. spindle assembly checkpoint [SAC]). A key event in this regard is the dephosphorylation of MELT repeats on KNL1, which removes SAC proteins from the kinetochore, including the BUB complex. We show here that PP1 and PP2A-B56 phosphatases are primarily required to remove Polo-like kinase 1 (PLK1) from the BUB complex, which can otherwise maintain MELT phosphorylation in an autocatalytic manner. This appears to be their principal role in the SAC because both phosphatases become redundant if PLK1 is inhibited or BUB-PLK1 interaction is prevented. Surprisingly, MELT dephosphorylation can occur normally under these conditions even when the levels or activities of PP1 and PP2A are strongly inhibited at kinetochores. Therefore, these data imply that kinetochore phosphatase regulation is critical for the SAC, but primarily to restrain and extinguish autonomous PLK1 activity. This is likely a conserved feature of the metazoan SAC, since the relevant PLK1 and PP2A-B56 binding motifs have coevolved in the same region on MADBUB homologues.
动粒处需要局部磷酸酶调节来沉默有丝分裂检验点(又称纺锤体组装检验点[SAC])。在这方面的一个关键事件是 KNL1 上 MELT 重复的去磷酸化,这会将 SAC 蛋白从动粒上移除,包括 BUB 复合物。我们在此表明,PP1 和 PP2A-B56 磷酸酶主要用于将 Polo 样激酶 1(PLK1)从 BUB 复合物上移除,否则 BUB 复合物可以以自催化的方式维持 MELT 的磷酸化。这似乎是它们在 SAC 中的主要作用,因为如果抑制 PLK1 或阻止 BUB-PLK1 相互作用,两种磷酸酶都会变得冗余。令人惊讶的是,即使在动粒处强烈抑制 PP1 和 PP2A 的水平或活性的情况下,MELT 的去磷酸化仍能正常发生。因此,这些数据表明动粒处的磷酸酶调节对于 SAC 至关重要,但主要是为了抑制和熄灭自主的 PLK1 活性。这可能是后生动物 SAC 的一个保守特征,因为相关的 PLK1 和 PP2A-B56 结合基序在 MADBUB 同源物上的同一区域共同进化。