Faculty of Electrical Engineering, University of Ljubljana, SI-1000 Ljubljana, Slovenia.
Faculty of Health Sciences, University of Ljubljana, SI-1000 Ljubljana, Slovenia.
Int J Mol Sci. 2021 Feb 26;22(5):2348. doi: 10.3390/ijms22052348.
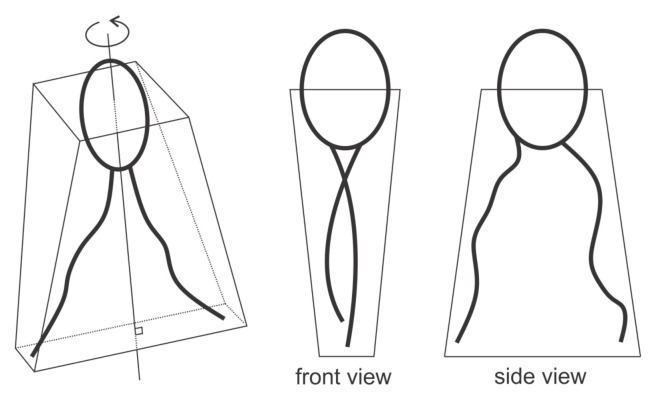
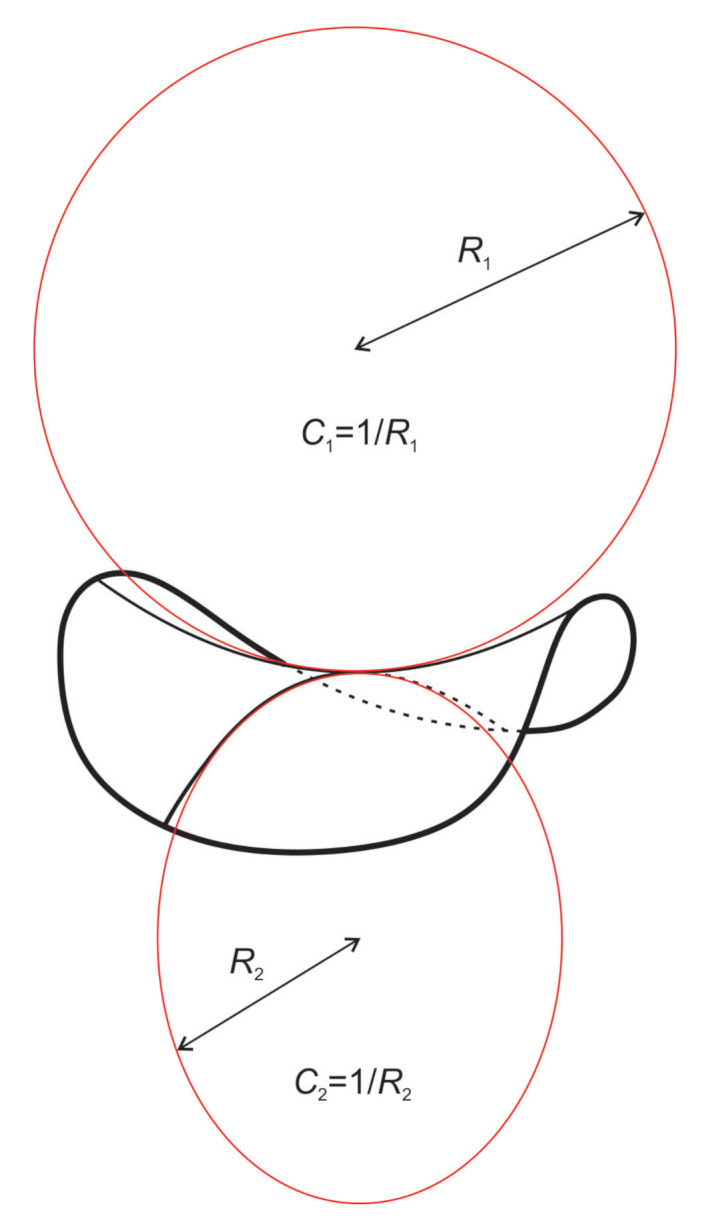
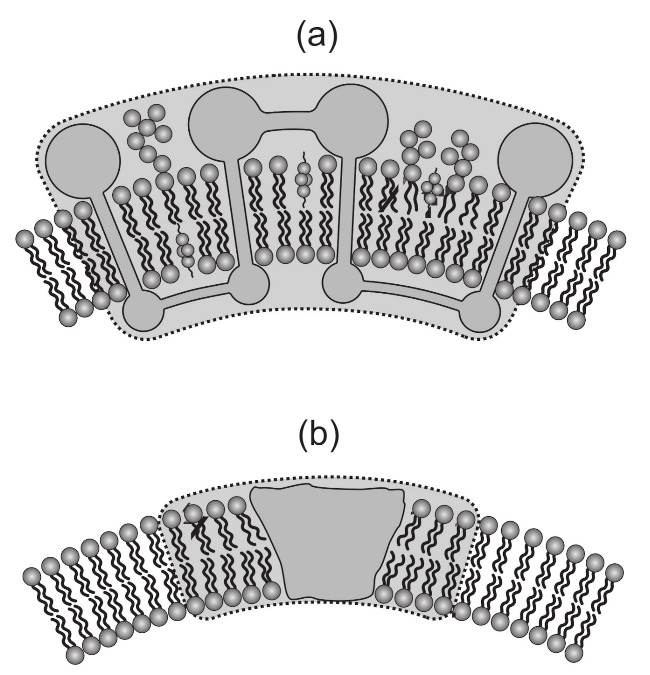
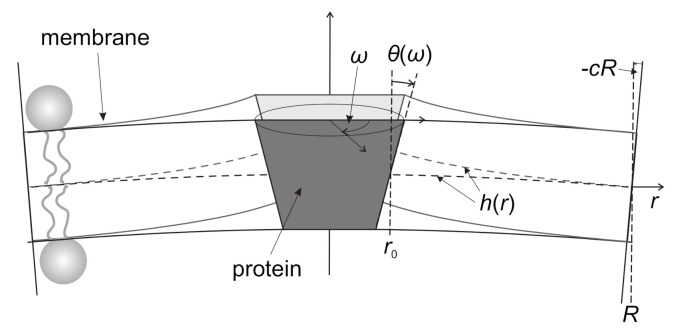
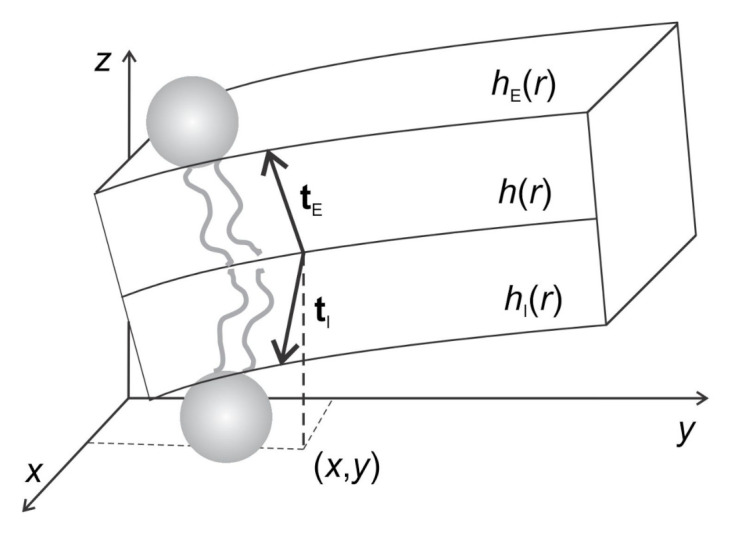
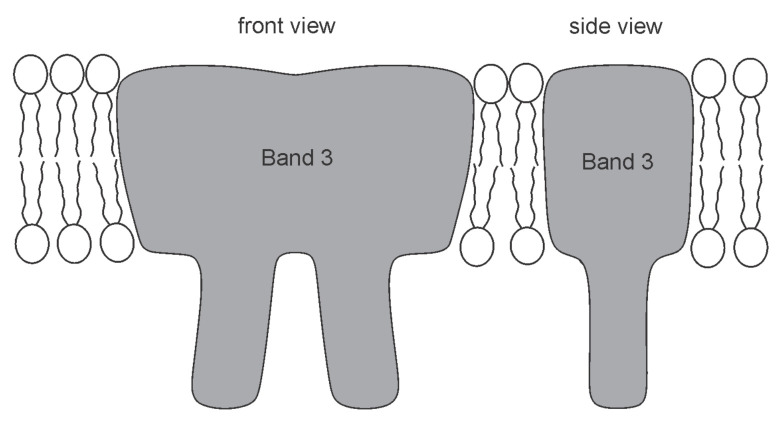
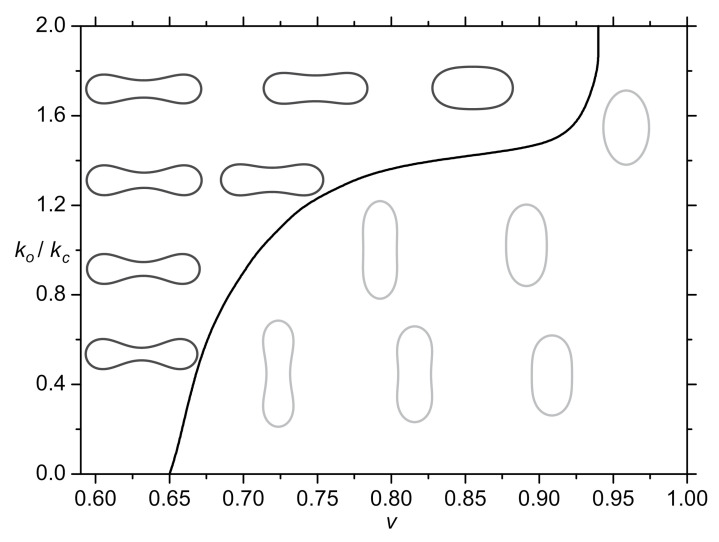
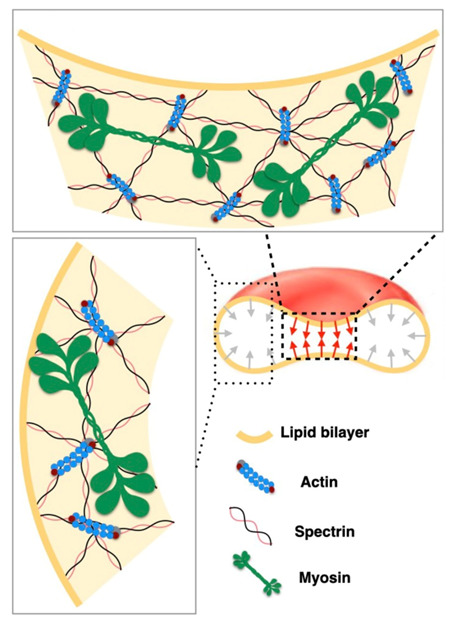
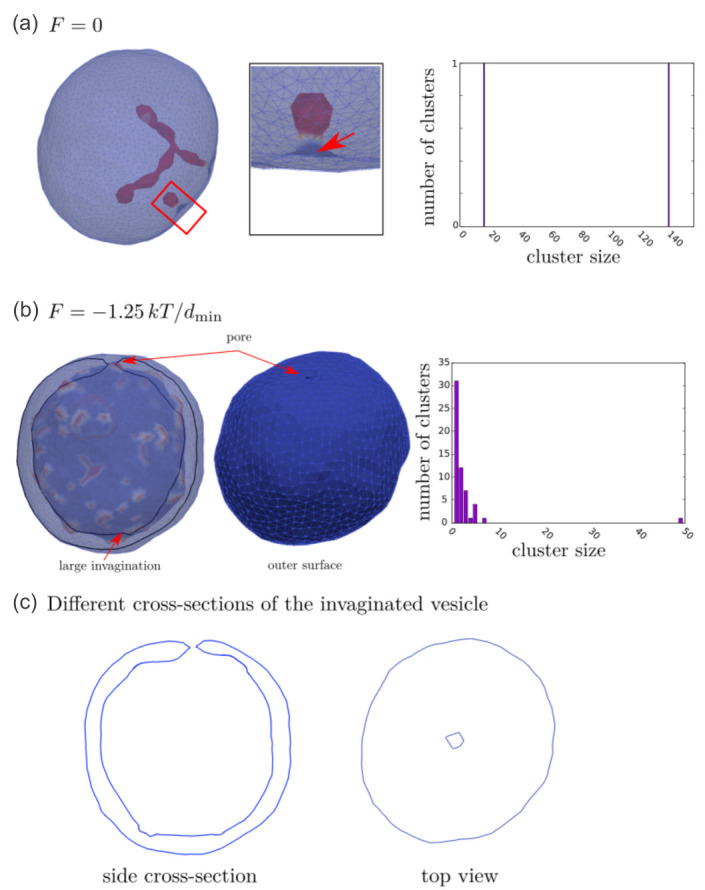
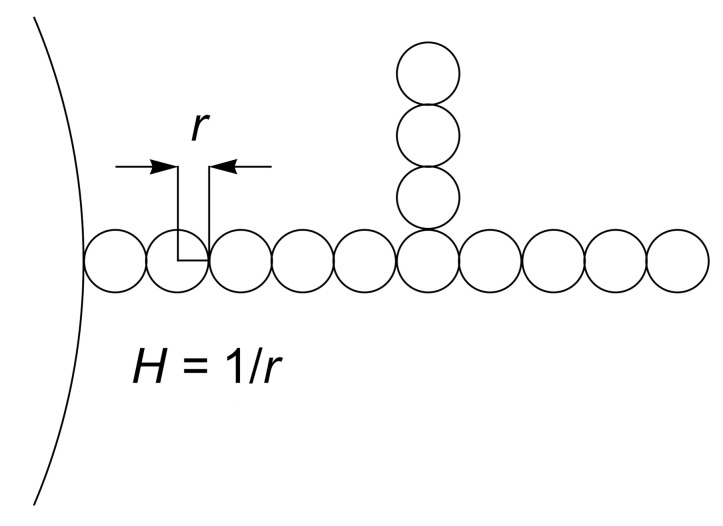
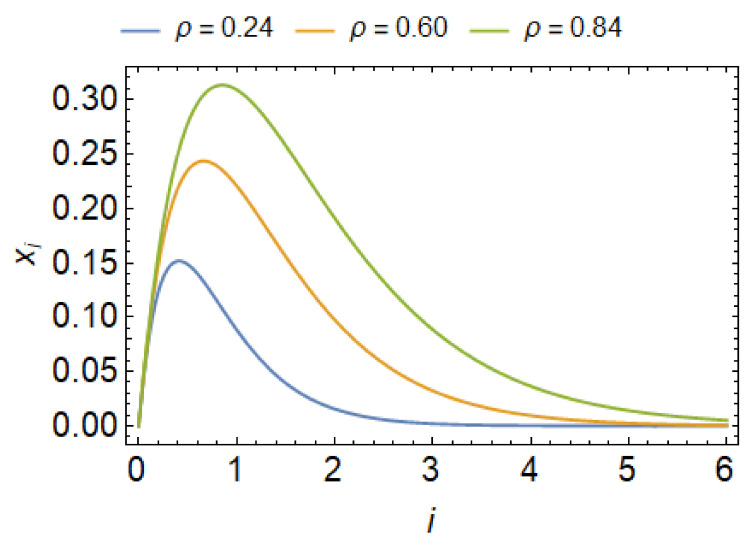
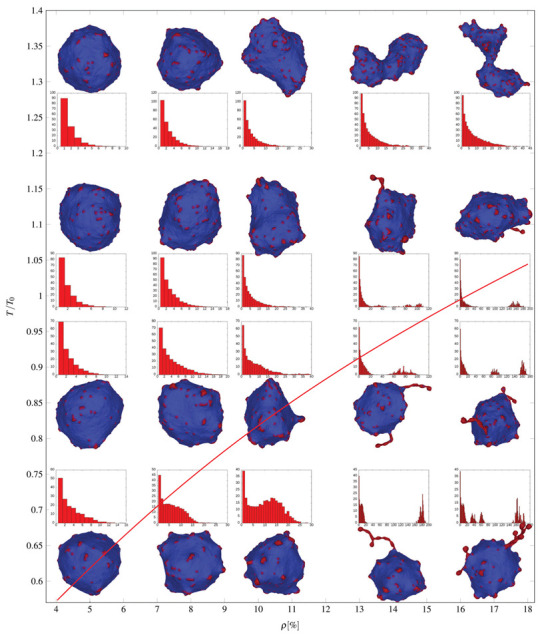
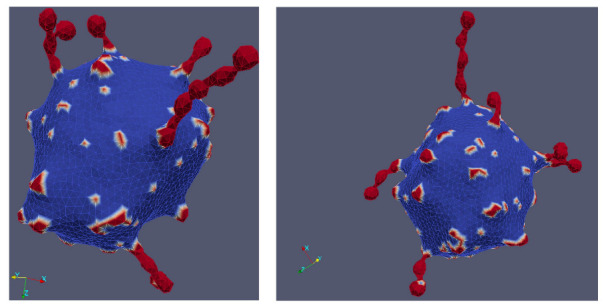
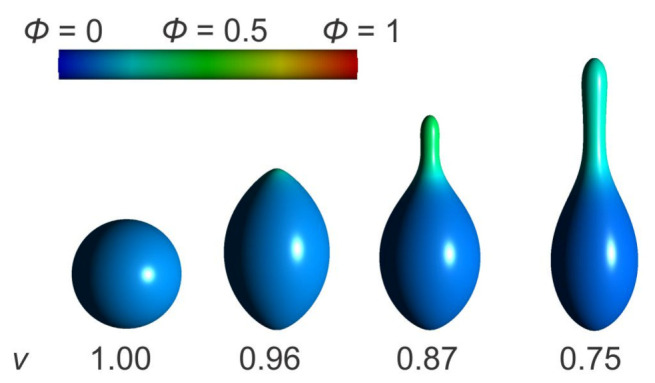
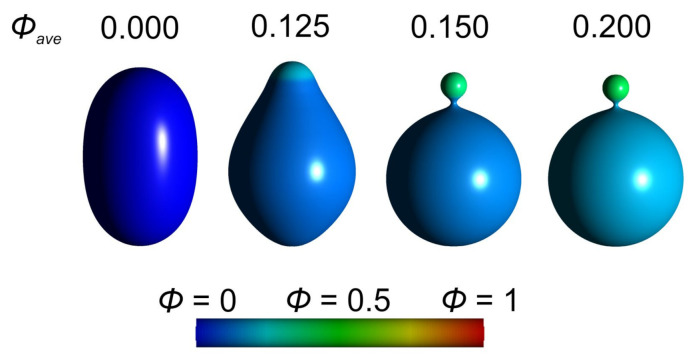
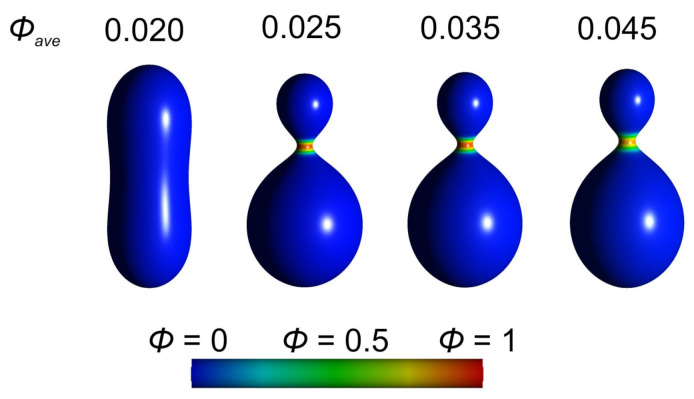
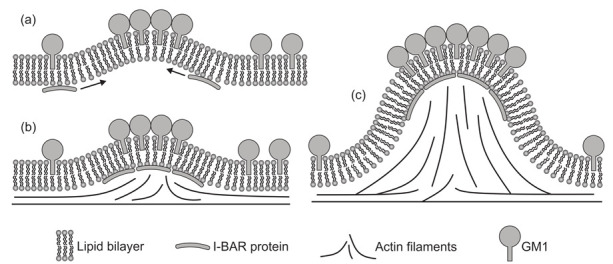
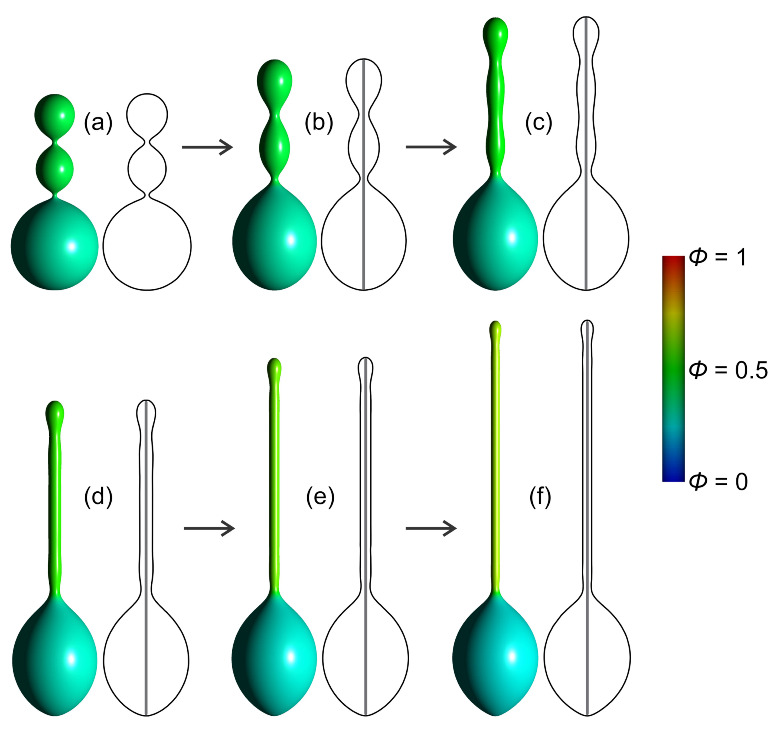
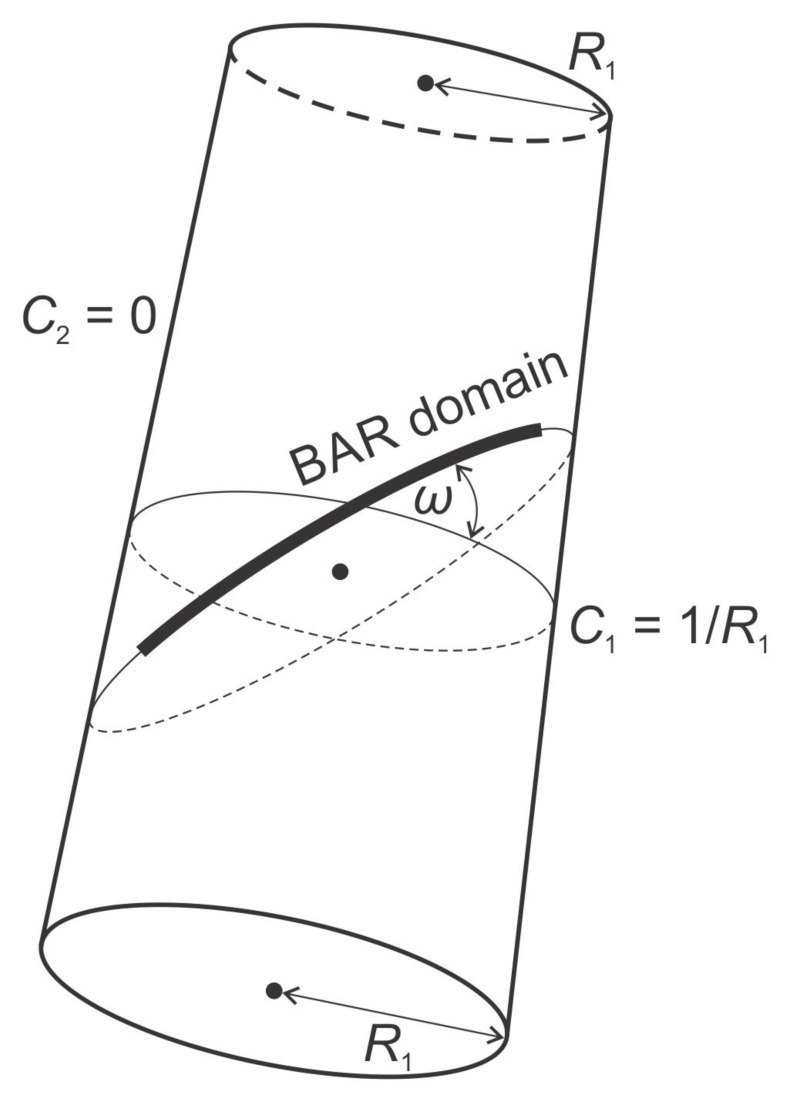
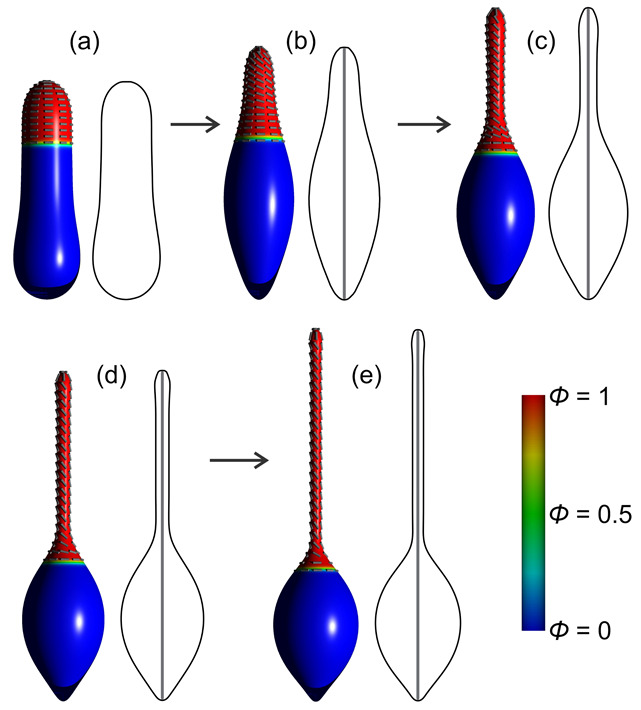
Biological membranes are composed of isotropic and anisotropic curved nanodomains. Anisotropic membrane components, such as Bin/Amphiphysin/Rvs (BAR) superfamily protein domains, could trigger/facilitate the growth of membrane tubular protrusions, while isotropic curved nanodomains may induce undulated (necklace-like) membrane protrusions. We review the role of isotropic and anisotropic membrane nanodomains in stability of tubular and undulated membrane structures generated or stabilized by cyto- or membrane-skeleton. We also describe the theory of spontaneous self-assembly of isotropic curved membrane nanodomains and derive the critical concentration above which the spontaneous necklace-like membrane protrusion growth is favorable. We show that the actin cytoskeleton growth inside the vesicle or cell can change its equilibrium shape, induce higher degree of segregation of membrane nanodomains or even alter the average orientation angle of anisotropic nanodomains such as BAR domains. These effects may indicate whether the actin cytoskeleton role is only to stabilize membrane protrusions or to generate them by stretching the vesicle membrane. Furthermore, we demonstrate that by taking into account the in-plane orientational ordering of anisotropic membrane nanodomains, direct interactions between them and the extrinsic (deviatoric) curvature elasticity, it is possible to explain the experimentally observed stability of oblate (discocyte) shapes of red blood cells in a broad interval of cell reduced volume. Finally, we present results of numerical calculations and Monte-Carlo simulations which indicate that the active forces of membrane skeleton and cytoskeleton applied to plasma membrane may considerably influence cell shape and membrane budding.
生物膜由各向同性和各向异性的弯曲纳米域组成。各向异性的膜成分,如 Bin/Amphiphysin/Rvs (BAR) 超家族蛋白域,可能触发/促进膜管状突起的生长,而各向同性弯曲纳米域可能诱导起皱(珠串状)的膜突起。我们回顾了各向同性和各向异性膜纳米域在由细胞骨架或膜骨架生成或稳定的管状和起皱膜结构稳定性中的作用。我们还描述了各向同性弯曲膜纳米域自发自组装的理论,并推导出了自发珠串状膜突起生长有利的临界浓度。我们表明,囊泡或细胞内肌动蛋白细胞骨架的生长可以改变其平衡形状,诱导膜纳米域更高程度的分离,甚至改变 BAR 域等各向异性纳米域的平均取向角。这些效应可能表明肌动蛋白细胞骨架的作用仅是稳定膜突起还是通过拉伸囊泡膜来生成它们。此外,我们证明,通过考虑各向异性膜纳米域的面内取向有序性、它们之间的直接相互作用以及外在(偏量)曲率弹性,可以解释在广泛的细胞体积减小区间内观察到的红细胞扁形(盘状)形状的稳定性。最后,我们呈现了数值计算和蒙特卡罗模拟的结果,表明施加于质膜的膜骨架和细胞骨架的主动力可能会显著影响细胞形状和膜出芽。