Institute of Pharmacology of Natural Products and Clinical Pharmacology, Ulm University, Ulm, Germany.
Department of Pharmaceutical Sciences, University of Basel, Basel, Switzerland.
Front Immunol. 2021 Feb 25;12:615748. doi: 10.3389/fimmu.2021.615748. eCollection 2021.
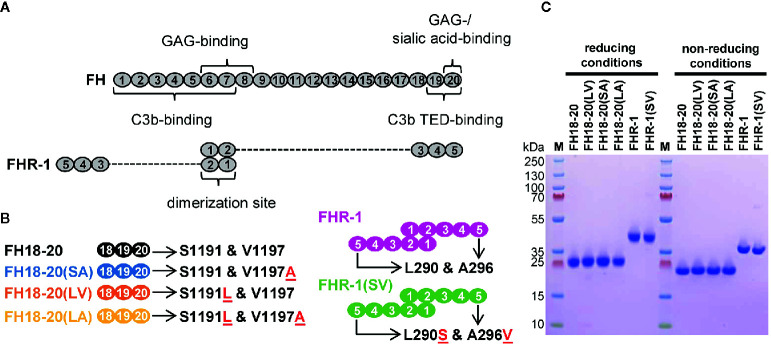
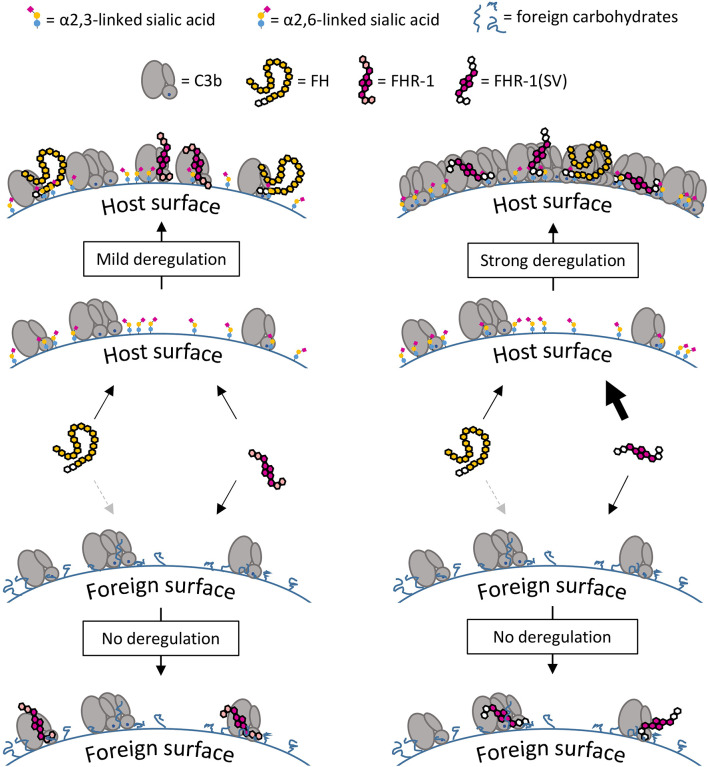
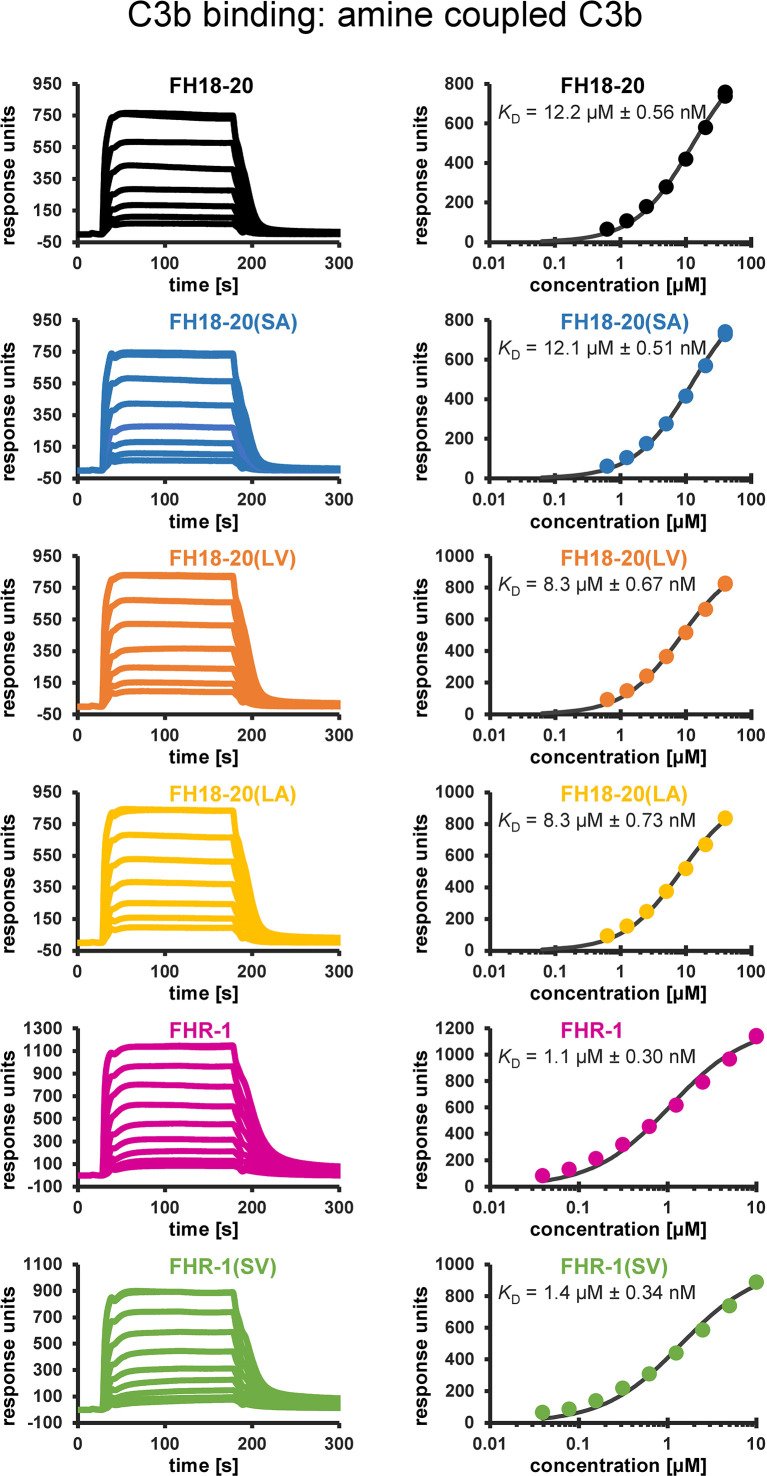
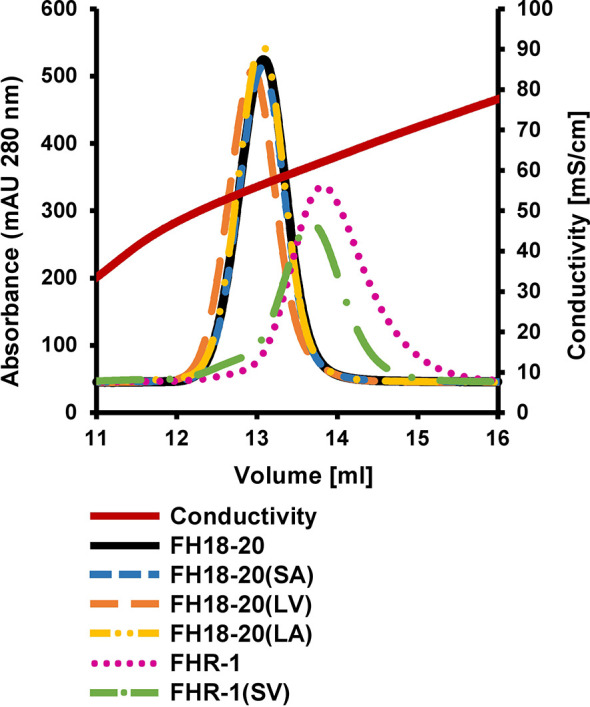
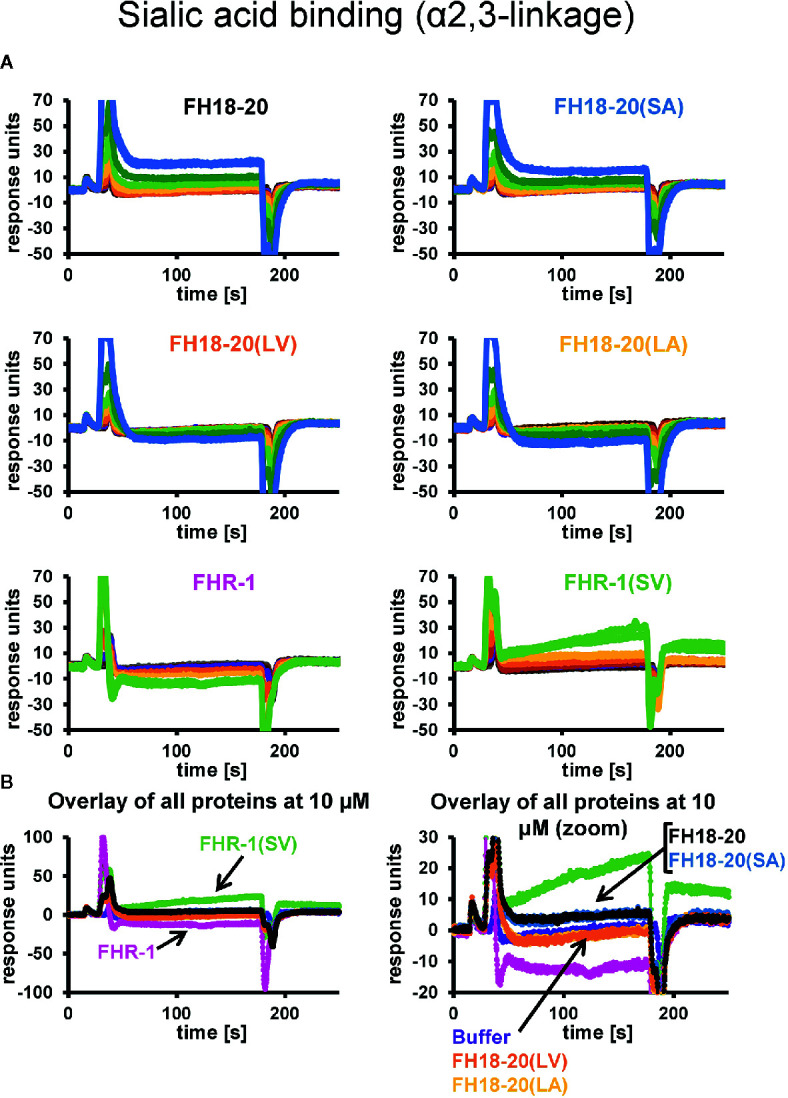
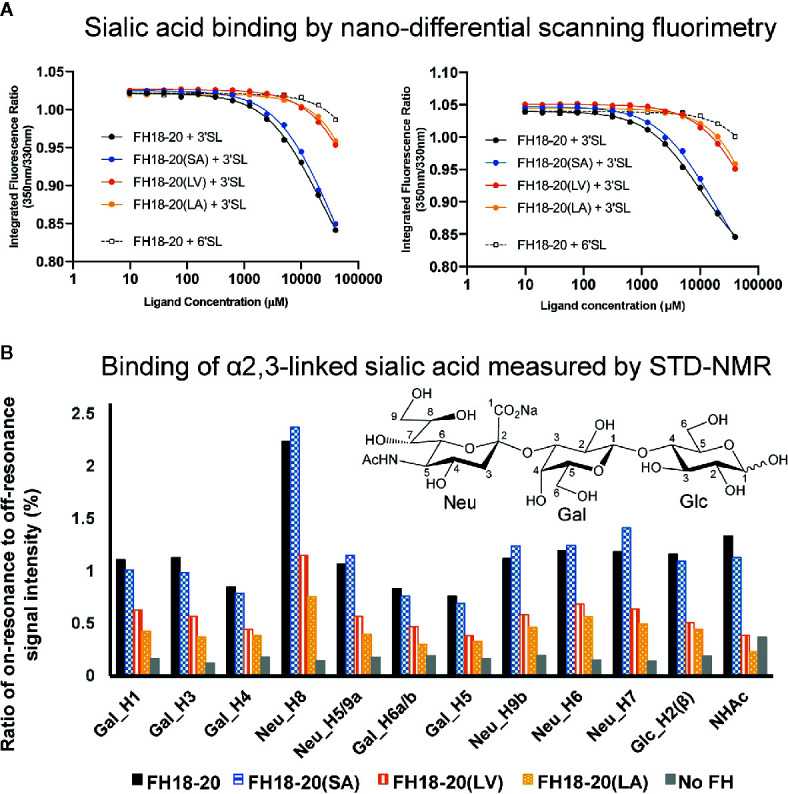
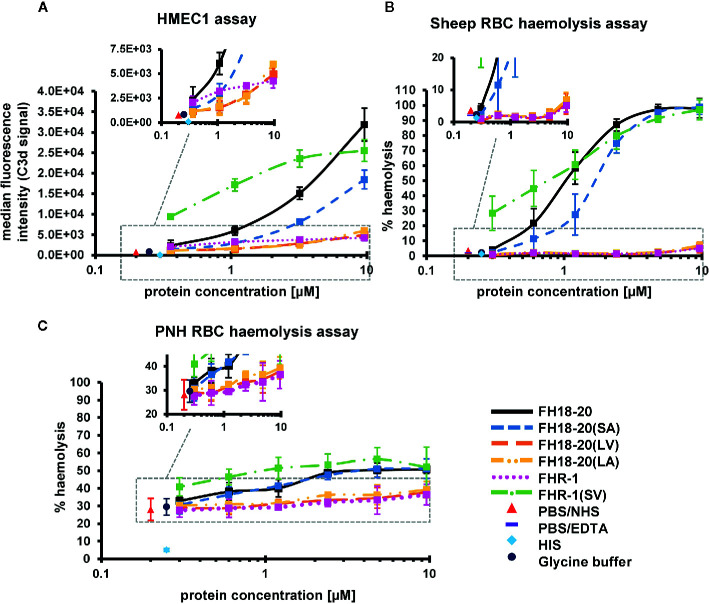
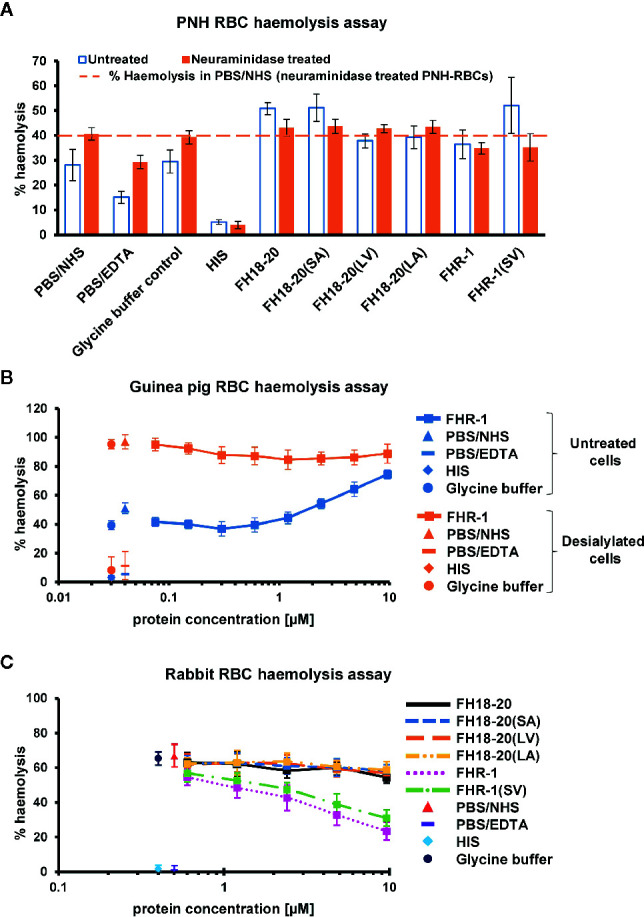
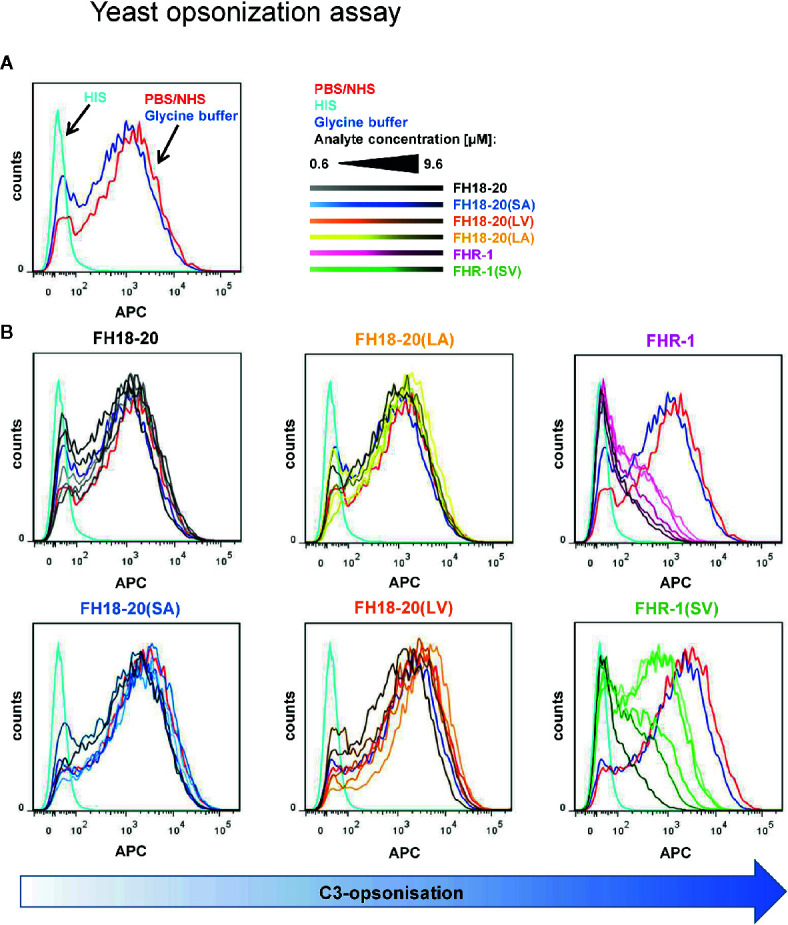
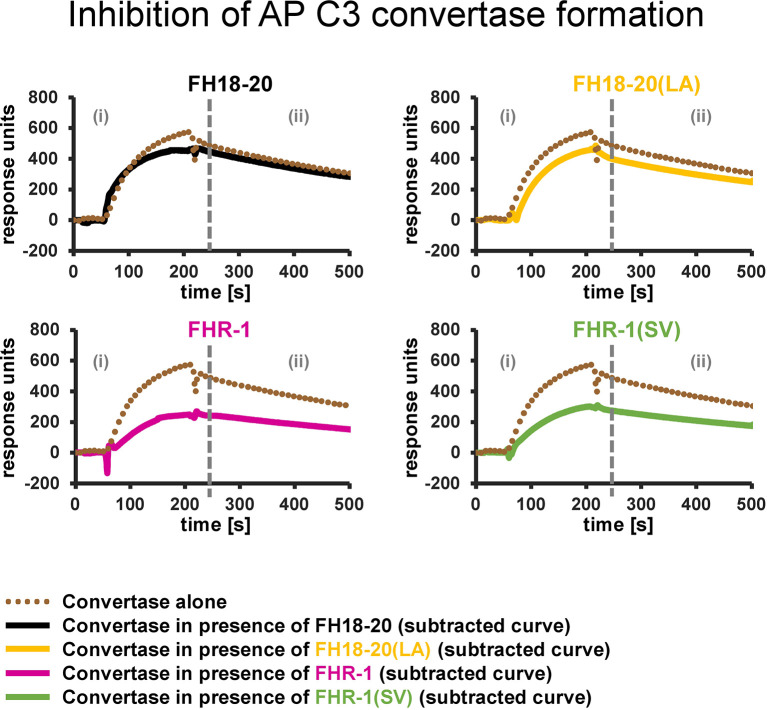
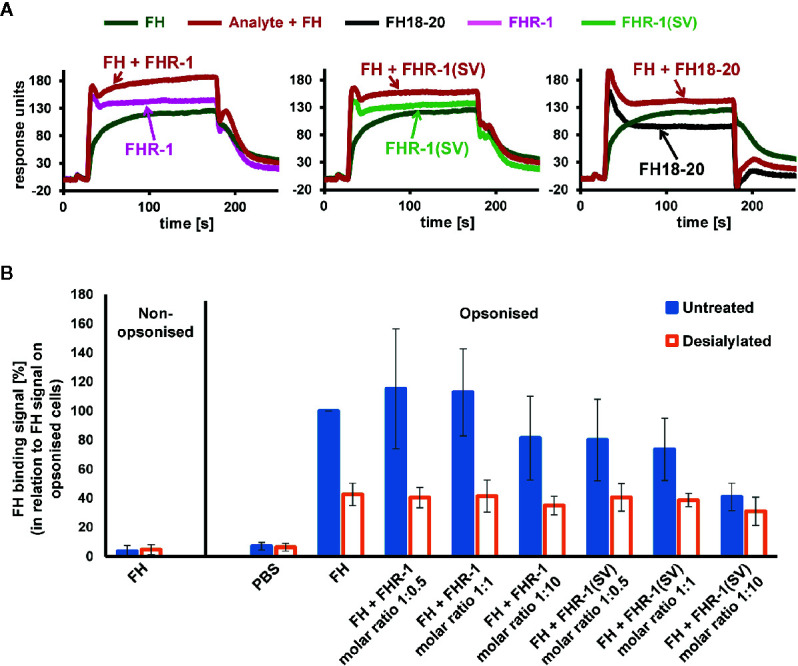
To discriminate between self and non-self surfaces and facilitate immune surveillance, the complement system relies on the interplay between surface-directed activators and regulators. The dimeric modulator FHR-1 is hypothesized to competitively remove the complement regulator FH from surfaces that strongly fix opsonic C3b molecules-a process known as "deregulation." The C-terminal regions of FH and FHR-1 provide the basis of this competition. They contain binding sites for C3b and host surface markers and are identical except for two substitutions: S1191L and V1197A (i.e., FH "SV"; FHR-1 "LA"). Intriguingly, an FHR-1 variant featuring the "SV" combination of FH predisposes to atypical hemolytic uremic syndrome (aHUS). The functional impact of these mutations on complement (de)regulation, and their pathophysiological consequences, have largely remained elusive. We have addressed these questions using recombinantly expressed wildtype, mutated, and truncated versions of FHR-1 and FH. The "SV" to "LA" substitutions did not affect glycosaminoglycan recognition and had only a small effect on C3b binding. In contrast, the two amino acids substantially affected the binding of FH and FHR-1 to α2,3-linked sialic acids as host surfaces markers, with the S-to-L substitution causing an almost complete loss of recognition. Even with sialic acid-binding constructs, notable deregulation was only detected on host and not foreign cells. The aHUS-associated "SV" mutation converts FHR-1 into a sialic acid binder which, supported by its dimeric nature, enables excessive FH deregulation and, thus, complement activation on host surfaces. While we also observed inhibitory activities of FHR-1 on C3 and C5 convertases, the high concentrations required render the physiological impact uncertain. In conclusion, the SV-to-LA substitution in the C-terminal regions of FH and FHR-1 diminishes its sialic acid-binding ability and results in an FHR-1 molecule that only moderately deregulates FH. Such FH deregulation by FHR-1 only occurs on host/host-like surfaces that recruit FH. Conversion of FHR-1 into a sialic acid binder potentiates the deregulatory capacity of FHR-1 and thus explains the pathophysiology of the aHUS-associated FHR-1 "SV" variant.
为了区分自身和非自身表面,并促进免疫监视,补体系统依赖于表面定向激活剂和调节剂之间的相互作用。二聚体调节剂 FHR-1 被假设通过竞争从强烈固定调理 C3b 分子的表面上除去补体调节剂 FH,这一过程称为“去调节”。FH 和 FHR-1 的 C 末端区域为这种竞争提供了基础。它们包含与 C3b 和宿主表面标记物结合的结合位点,并且除了两个取代之外是相同的:S1191L 和 V1197A(即 FH“SV”;FHR-1“LA”)。有趣的是,具有 FH 的“SV”组合的 FHR-1 变体易患非典型溶血性尿毒症综合征(aHUS)。这些突变对补体(去)调节的功能影响及其病理生理后果在很大程度上仍然难以捉摸。我们使用重组表达的野生型、突变型和截短型 FHR-1 和 FH 来解决这些问题。“SV”到“LA”取代不影响糖胺聚糖识别,仅对 C3b 结合有微小影响。相比之下,这两个氨基酸会显著影响 FH 和 FHR-1 与作为宿主表面标记物的α2,3 连接唾液酸的结合,其中 S 到 L 的取代导致几乎完全丧失识别。即使使用唾液酸结合构建体,也仅在宿主细胞而不是外来细胞上检测到明显的去调节。与 aHUS 相关的“SV”突变将 FHR-1 转化为唾液酸结合物,该结合物通过其二聚体性质,能够在宿主表面上过度调节 FH,并因此激活补体。虽然我们还观察到 FHR-1 对 C3 和 C5 转化酶的抑制活性,但所需的高浓度使其生理影响不确定。总之,FH 和 FHR-1 的 C 末端区域的 SV 到 LA 取代降低了其唾液酸结合能力,导致 FHR-1 分子仅适度地调节 FH。仅在招募 FH 的宿主/宿主样表面上才会发生 FHR-1 对 FH 的这种去调节。FHR-1 转化为唾液酸结合物增强了 FHR-1 的去调节能力,从而解释了与 aHUS 相关的 FHR-1“SV”变体的病理生理学。