Department of Biological Sciences, University of Pittsburgh, Pittsburgh, United States.
Women's Cancer Research Center, UPMC Hillman Cancer Center, Magee-Women Research Institute, Pittsburgh, United States.
Elife. 2021 Jun 28;10:e64977. doi: 10.7554/eLife.64977.
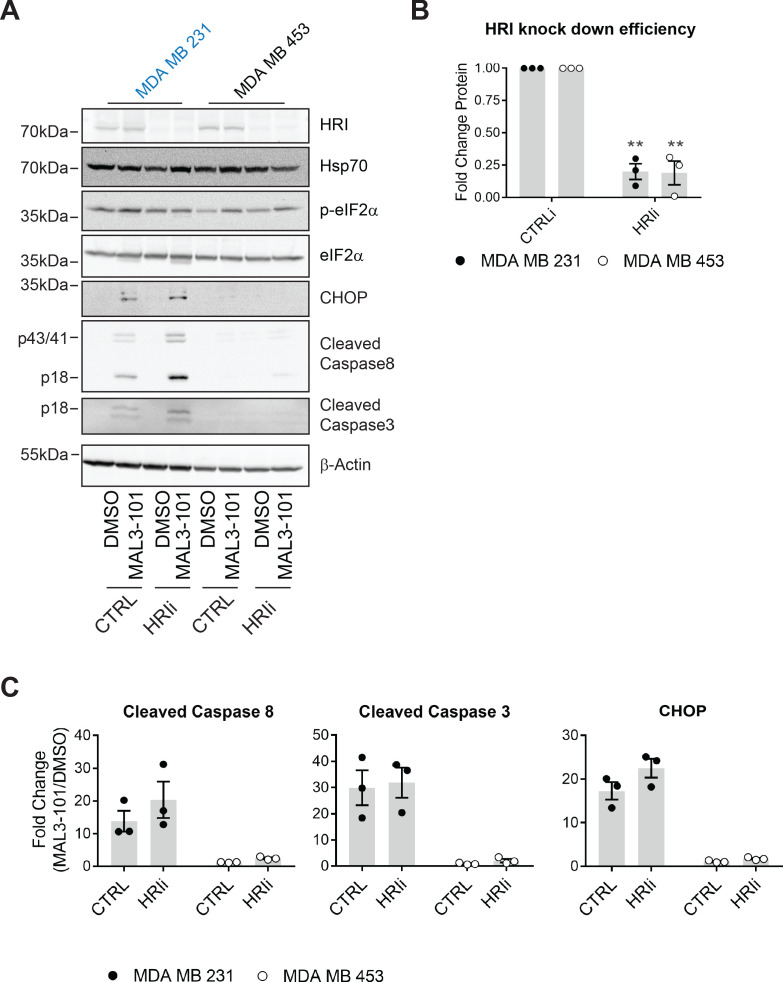
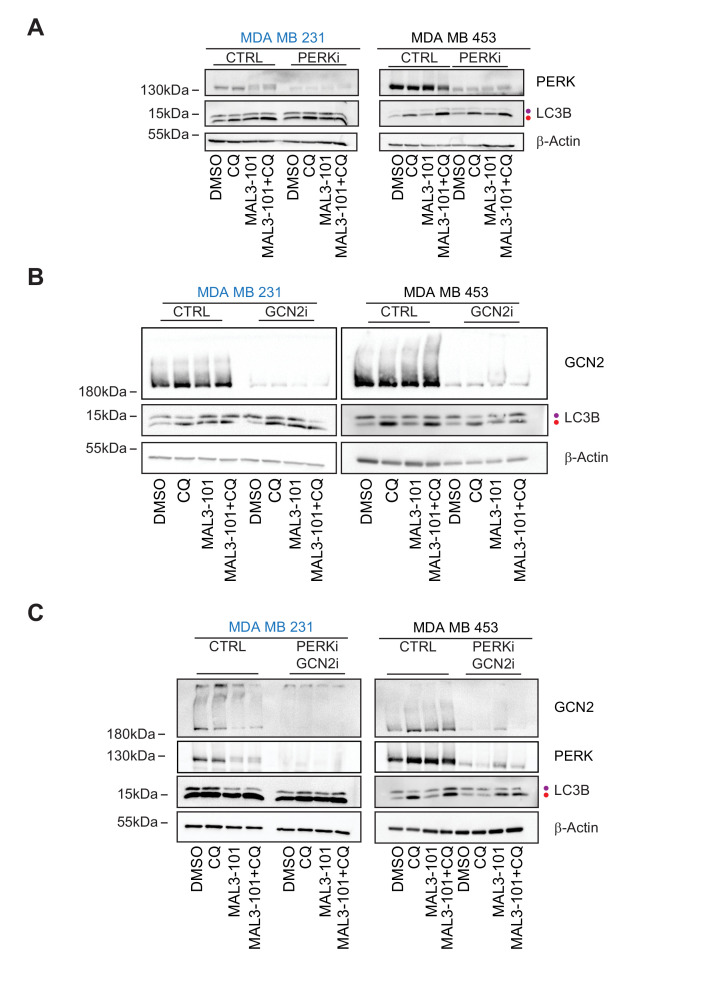
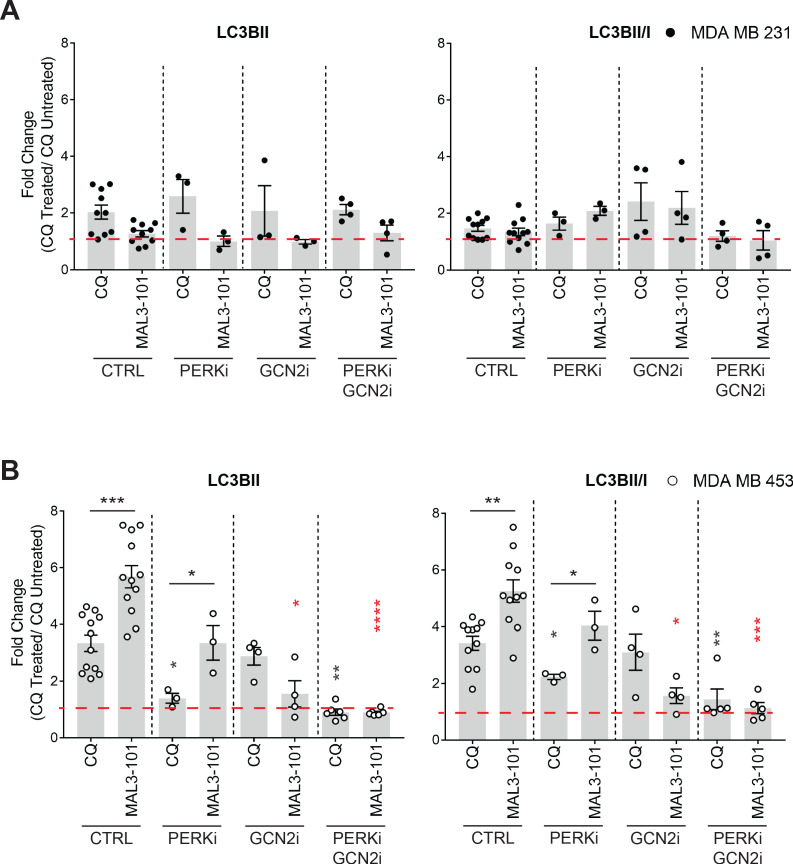
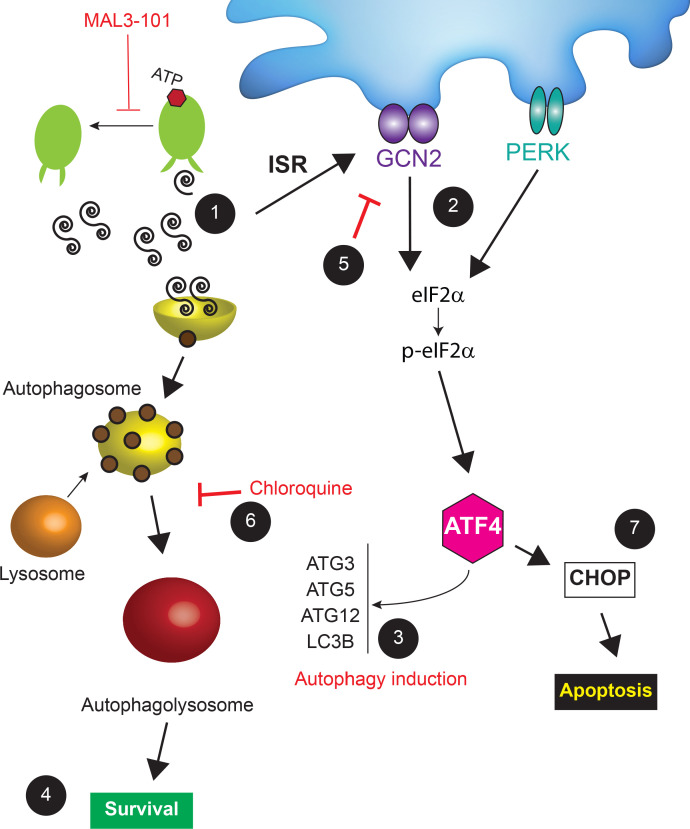
Molecular chaperones, such as Hsp70, prevent proteotoxicity and maintain homeostasis. This is perhaps most evident in cancer cells, which overexpress Hsp70 and thrive even when harboring high levels of misfolded proteins. To define the response to proteotoxic challenges, we examined adaptive responses in breast cancer cells in the presence of an Hsp70 inhibitor. We discovered that the cells bin into distinct classes based on inhibitor sensitivity. Strikingly, the most resistant cells have higher autophagy levels, and autophagy was maximally activated only in resistant cells upon Hsp70 inhibition. In turn, resistance to compromised Hsp70 function required the integrated stress response transducer, GCN2, which is commonly associated with amino acid starvation. In contrast, sensitive cells succumbed to Hsp70 inhibition by activating PERK. These data reveal an unexpected route through which breast cancer cells adapt to proteotoxic insults and position GCN2 and autophagy as complementary mechanisms to ensure survival when proteostasis is compromised.
分子伴侣,如 Hsp70,可以防止蛋白毒性并维持细胞内环境稳定。这在癌细胞中表现得最为明显,癌细胞过度表达 Hsp70,即使存在高水平的错误折叠蛋白,也能茁壮成长。为了定义对蛋白毒性挑战的反应,我们研究了在存在 Hsp70 抑制剂的情况下乳腺癌细胞的适应性反应。我们发现,根据抑制剂的敏感性,细胞可以分为不同的类别。引人注目的是,最具抗性的细胞具有更高的自噬水平,并且只有在抗性细胞中,Hsp70 抑制才会最大程度地激活自噬。反过来,对 Hsp70 功能受损的抗性需要整合应激反应转导物 GCN2,这通常与氨基酸饥饿有关。相比之下,敏感细胞通过激活 PERK 来对抗 Hsp70 抑制。这些数据揭示了乳腺癌细胞适应蛋白毒性损伤的一条意外途径,并将 GCN2 和自噬定位为互补机制,以在蛋白稳态受损时确保存活。