Laboratório de Bioquímica de Insetos, Instituto de Bioquímica Médica Leopoldo de Meis. Universidade Federal do Rio de Janeiro, Brazil.
Laboratório de Imunologia Molecular e Celular, Instituto de Biofísica Carlos Chagas Filho (IBCCF), Universidade Federal do Rio de Janeiro, Brazil.
PLoS Negl Trop Dis. 2021 Sep 7;15(9):e0009760. doi: 10.1371/journal.pntd.0009760. eCollection 2021 Sep.
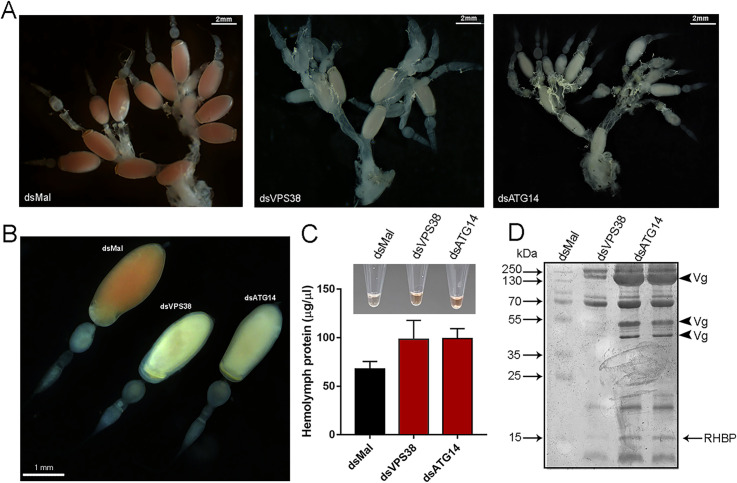
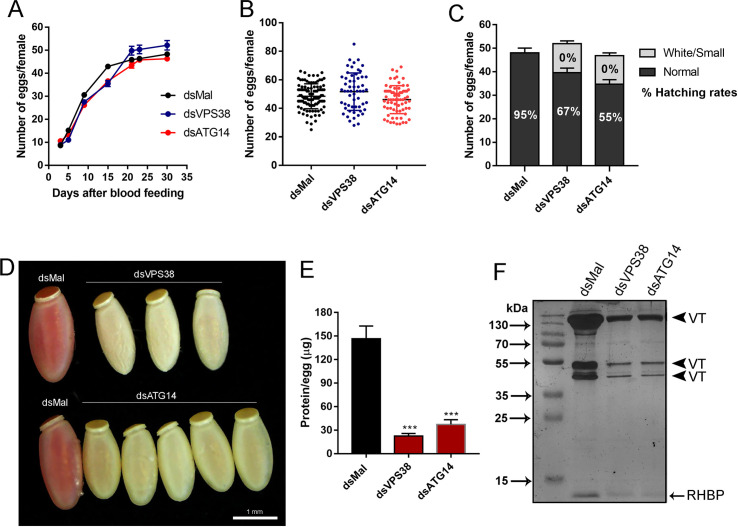
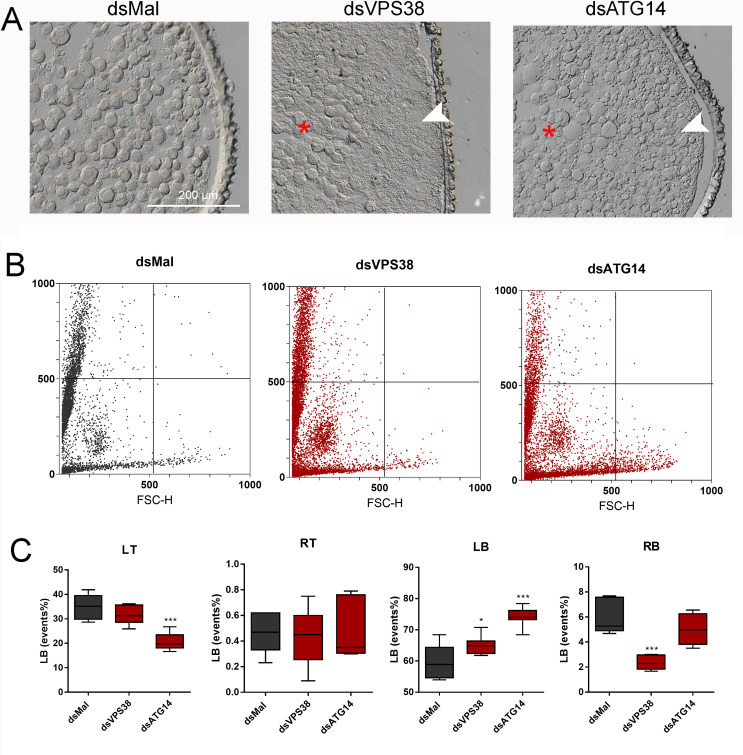
In insects the reserve proteins are stored in the oocytes into endocytic-originated vesicles named yolk organelles. VPS38/UVRAG and ATG14 are the variant regulatory subunits of two class-III ATG6/Beclin1 PI3K complexes that regulate the recruitment of the endocytic (complex II) and autophagic (complex I) machineries. In a previous work from our group, we found that the silencing of ATG6/Beclin1 resulted in the formation of yolk-deficient oocytes due to defects in the endocytosis of the yolk proteins. Because ATG6/Beclin1 is present in the two above-described PI3K complexes, we could not identify the contributions of each complex to the yolk defective phenotypes. To address this, here we investigated the role of the variant subunits VPS38/UVRAG (complex II, endocytosis) and ATG14 (complex I, autophagy) in the biogenesis of the yolk organelles in the insect vector of Chagas Disease Rhodnius prolixus. Interestingly, the silencing of both genes phenocopied the silencing of ATG6/Beclin1, generating 1) accumulation of yolk proteins in the hemolymph; 2) white, smaller, and yolk-deficient oocytes; 3) abnormal yolk organelles in the oocyte cortex; and 4) unviable F1 embryos. However, we found that the similar phenotypes were the result of a specific cross-silencing effect among the PI3K subunits where the silencing of VPS38/UVRAG and ATG6/Beclin1 resulted in the specific silencing of each other, whereas the silencing of ATG14 triggered the silencing of all three PI3K components. Because the silencing of VPS38/UVRAG and ATG6/Beclin1 reproduced the yolk-deficiency phenotypes without the cross silencing of ATG14, we concluded that the VPS38/UVRAG PI3K complex II was the major contributor to the previously observed phenotypes in silenced insects. Altogether, we found that class-III ATG6/Beclin1 PI3K complex II (VPS38/UVRAG) is essential for the yolk endocytosis and that the subunits of both complexes are under an unknown transcriptional regulatory system.
在昆虫中,储备蛋白储存在卵母细胞中,形成内吞起源的小泡,称为卵黄细胞器。VPS38/UVRAG 和 ATG14 是两种 III 类 ATG6/Beclin1 PI3K 复合物的变构调节亚基,它们调节内吞(复合物 II)和自噬(复合物 I)机器的募集。在我们之前的工作中,我们发现 ATG6/Beclin1 的沉默导致卵黄蛋白内吞缺陷,从而导致卵黄缺乏的卵母细胞形成。因为 ATG6/Beclin1 存在于上述两种 PI3K 复合物中,我们无法确定每个复合物对卵黄缺陷表型的贡献。为了解决这个问题,我们在这里研究了变构亚基 VPS38/UVRAG(复合物 II,内吞作用)和 ATG14(复合物 I,自噬作用)在恰加斯病传播媒介 Rhodnius prolixus 中卵黄细胞器生物发生中的作用。有趣的是,这两个基因的沉默表型与 ATG6/Beclin1 的沉默表型相似,导致 1)卵黄蛋白在血淋巴中积累;2)白色、较小和卵黄缺乏的卵母细胞;3)卵母细胞皮质中的异常卵黄细胞器;4)无法存活的 F1 胚胎。然而,我们发现,相似的表型是 PI3K 亚基之间特定的交叉沉默效应的结果,其中 VPS38/UVRAG 和 ATG6/Beclin1 的沉默导致彼此的特异性沉默,而 ATG14 的沉默则触发了所有三个 PI3K 成分的沉默。由于 VPS38/UVRAG 和 ATG6/Beclin1 的沉默复制了卵黄缺乏的表型,而没有 ATG14 的交叉沉默,我们得出结论,VPS38/UVRAG PI3K 复合物 II 是先前在沉默昆虫中观察到的表型的主要贡献者。总的来说,我们发现 III 类 ATG6/Beclin1 PI3K 复合物 II(VPS38/UVRAG)对于卵黄内吞作用是必不可少的,并且两个复合物的亚基都受到未知的转录调控系统的调节。