Lambey Paul, Otun Omolade, Cong Xiaojing, Hoh François, Brunel Luc, Verdié Pascal, Grison Claire M, Peysson Fanny, Jeannot Sylvain, Durroux Thierry, Bechara Cherine, Granier Sébastien, Leyrat Cédric
Institut de Génomique Fonctionnelle, Université de Montpellier, CNRS, INSERM, Montpellier, France.
Centre de Biochimie Structurale, CNRS UMR 5048-INSERM 1054- University of Montpellier, Montpellier, France.
Elife. 2022 Mar 21;11:e72555. doi: 10.7554/eLife.72555.
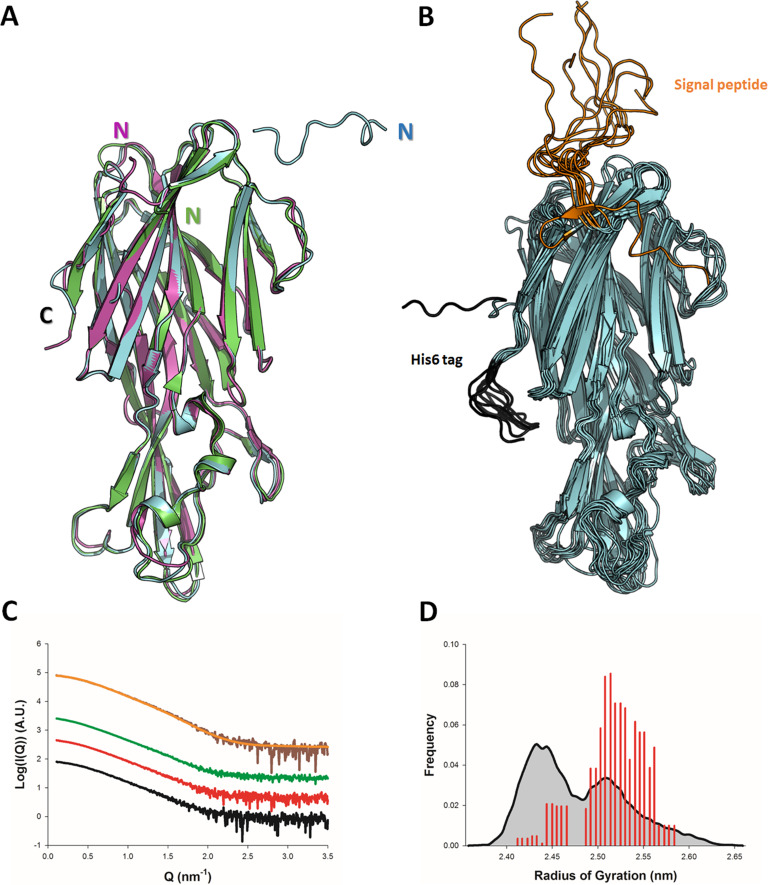
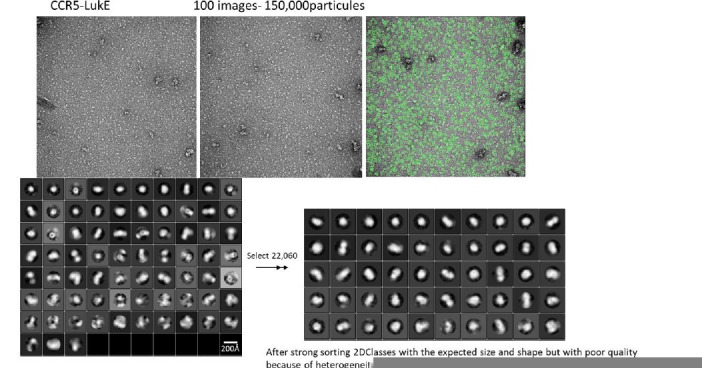
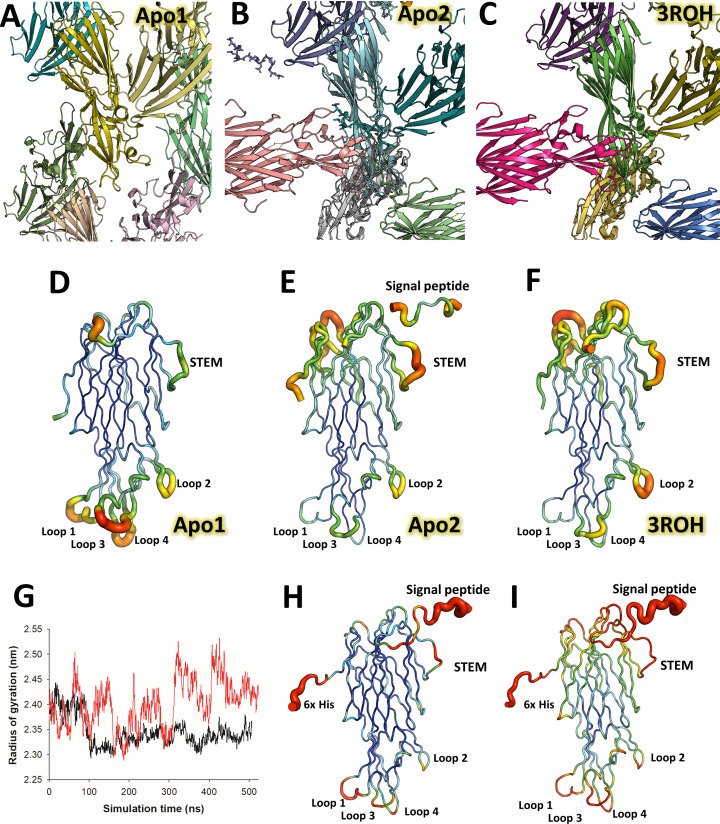
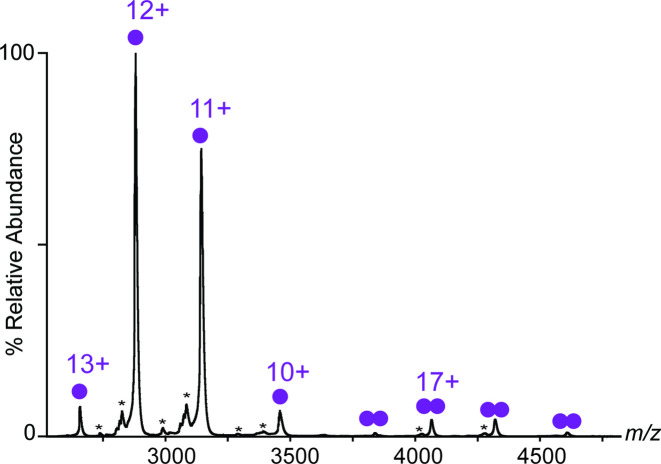
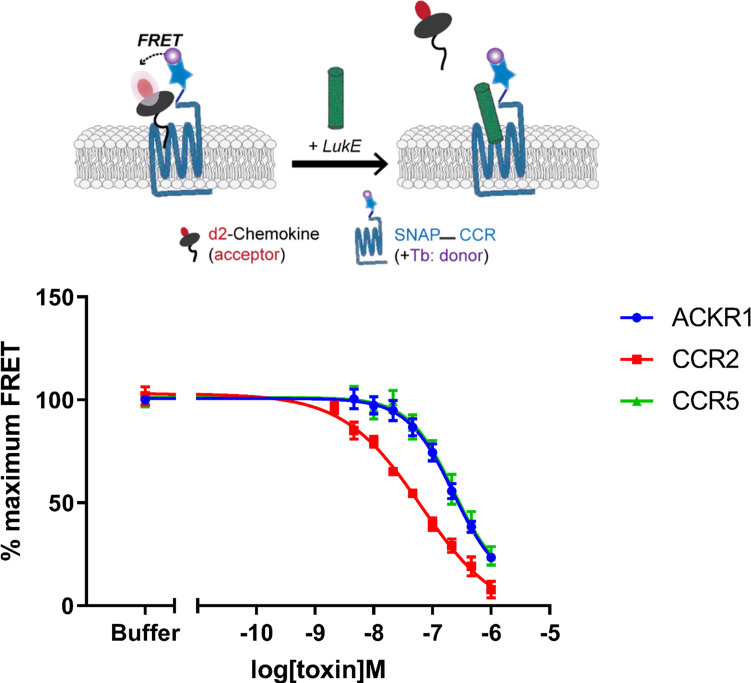
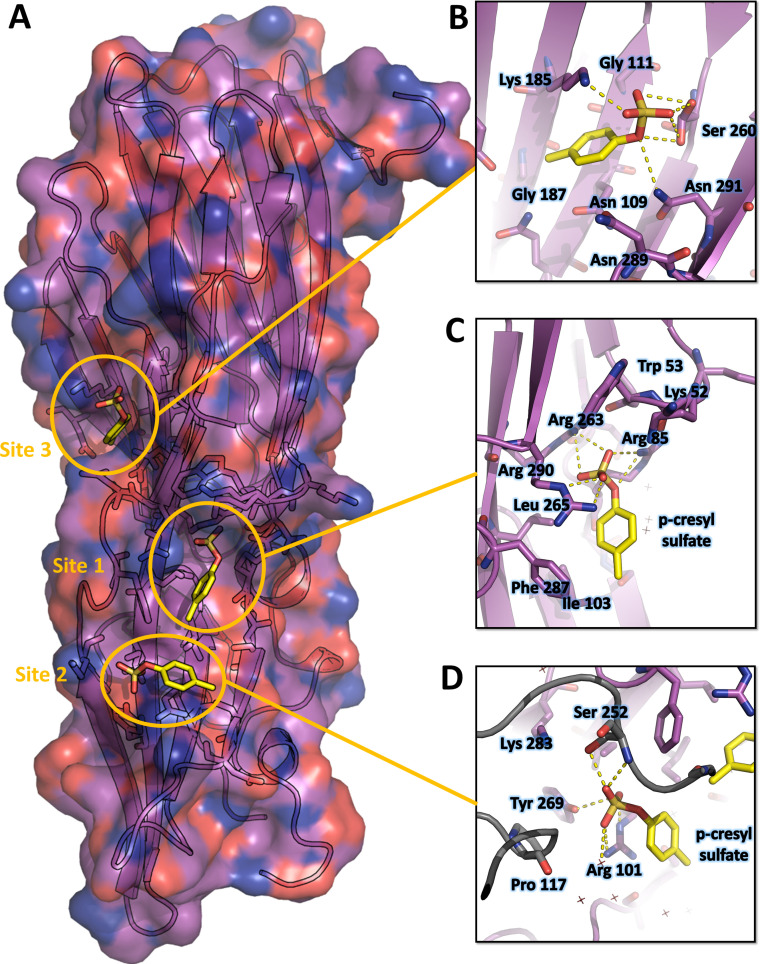
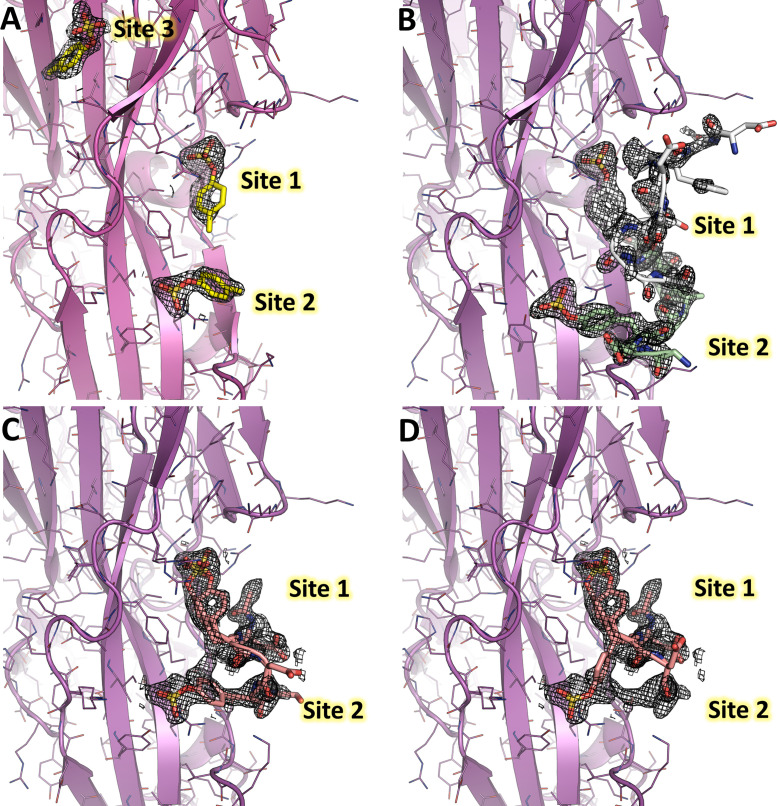
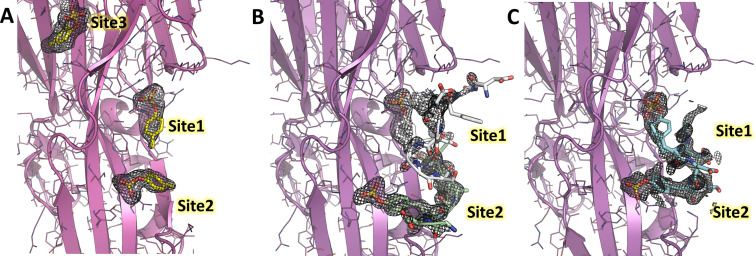
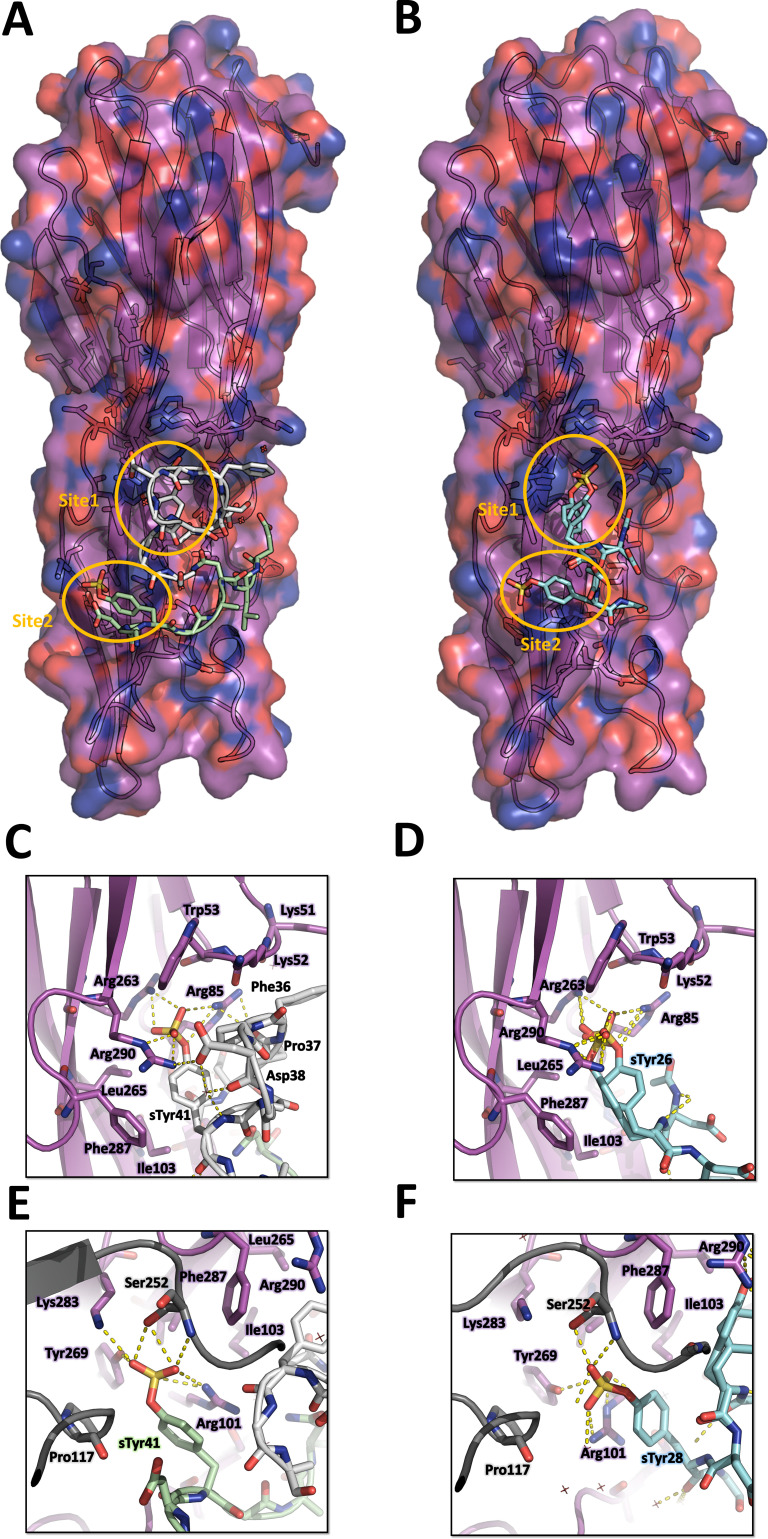
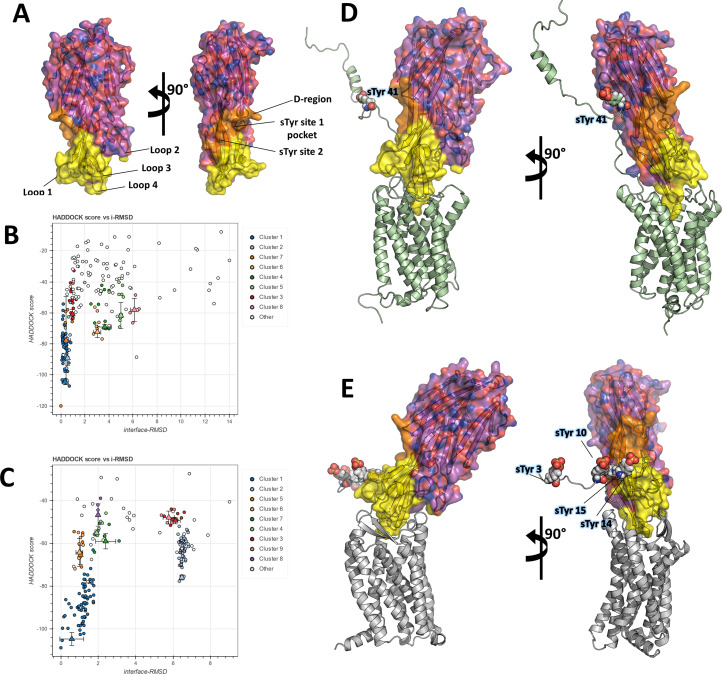
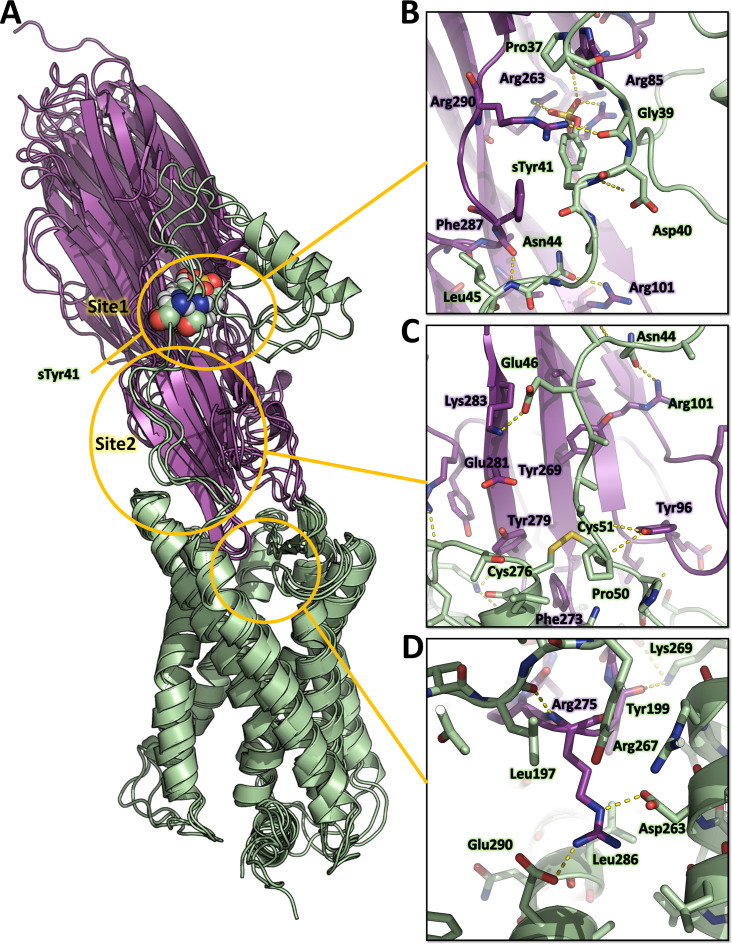
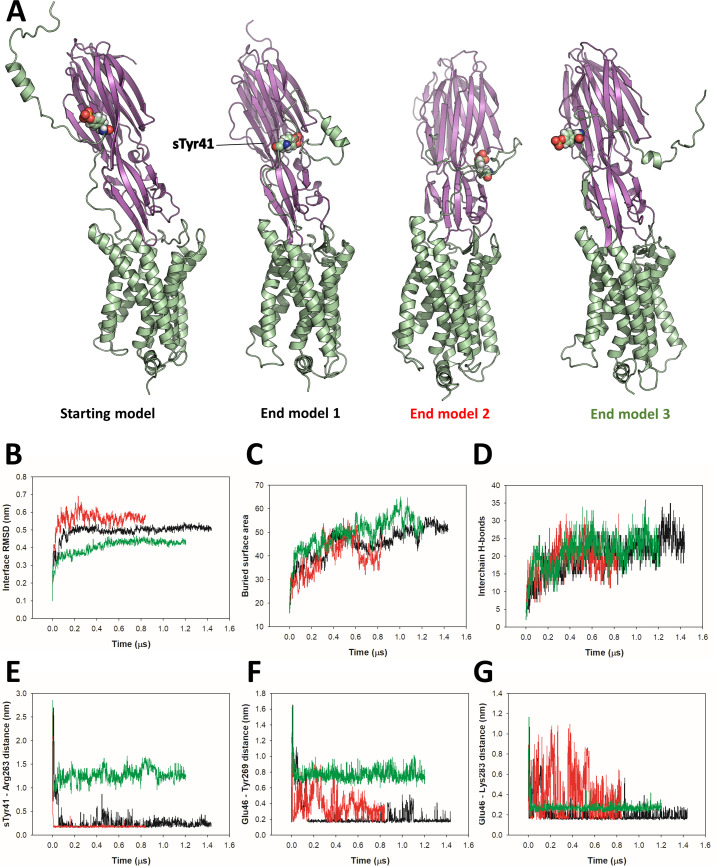
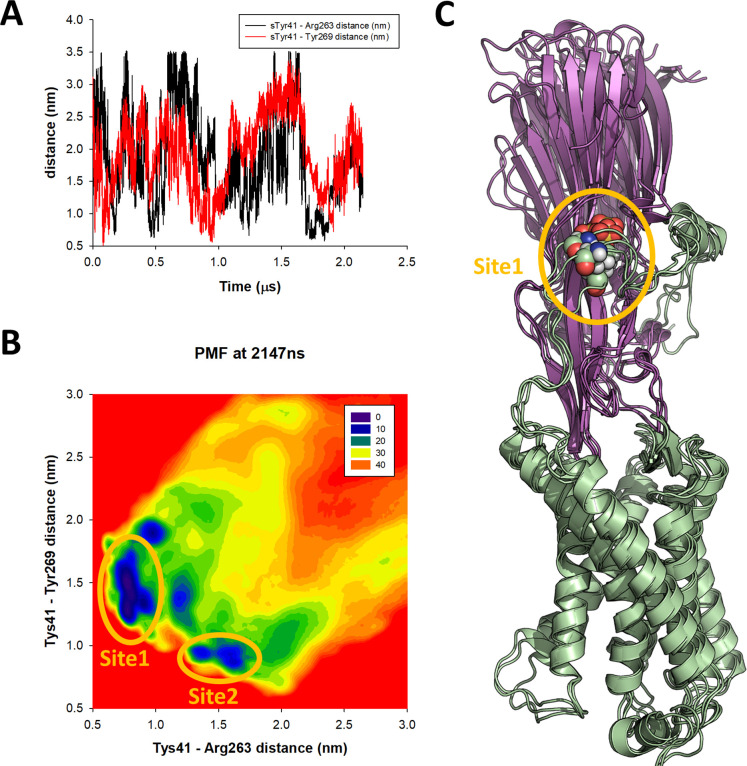
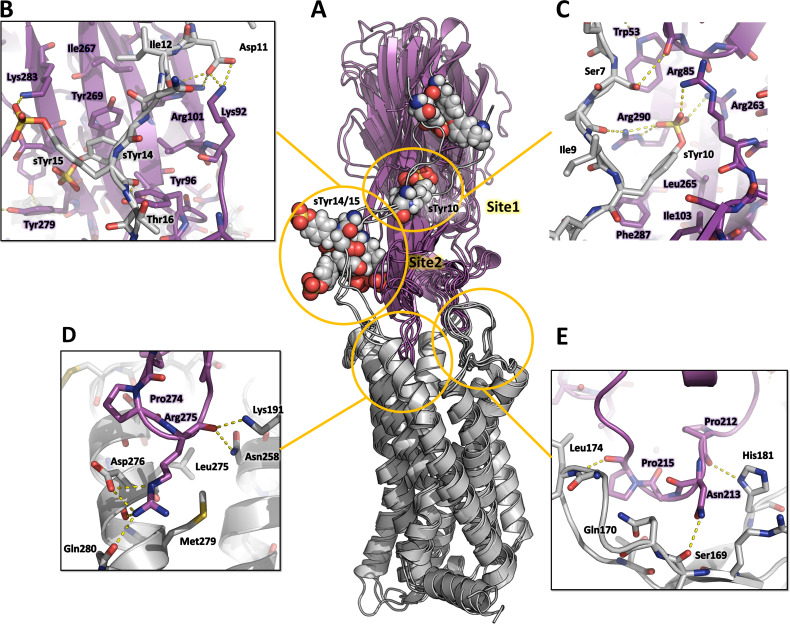
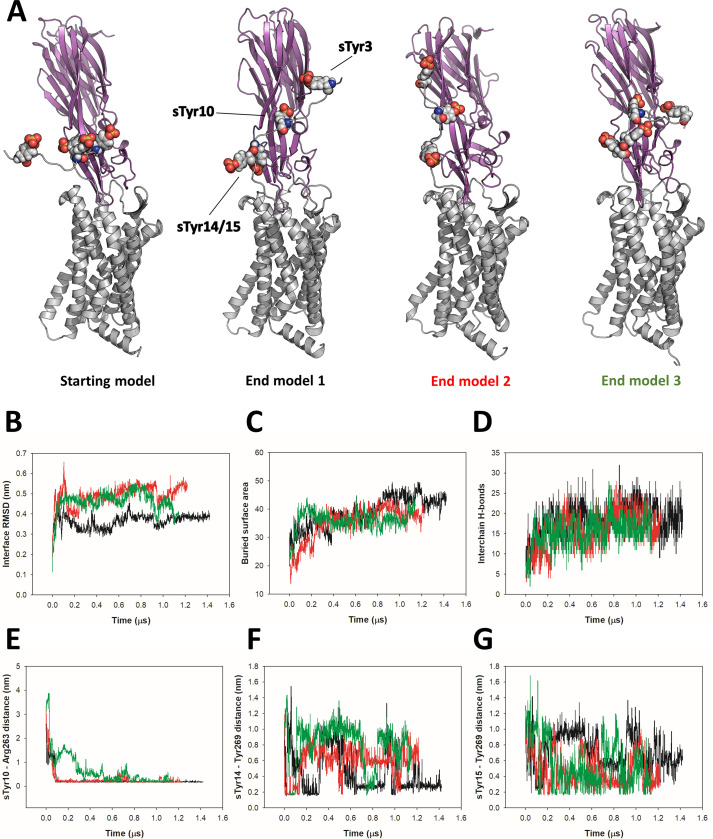
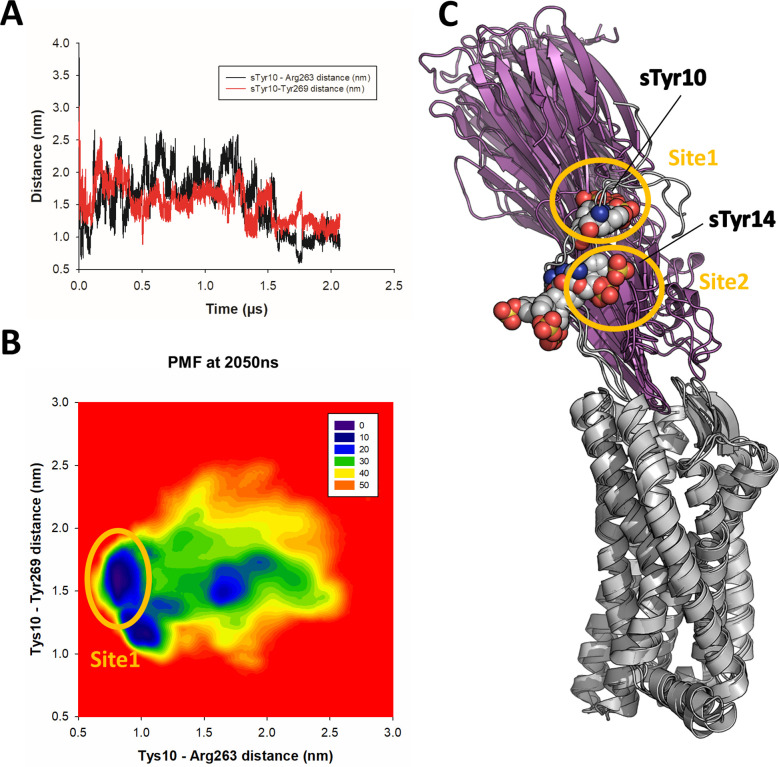
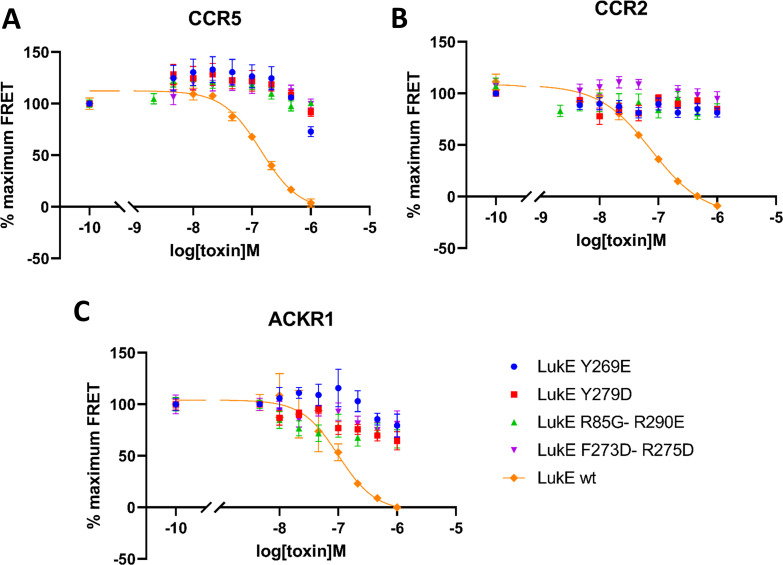
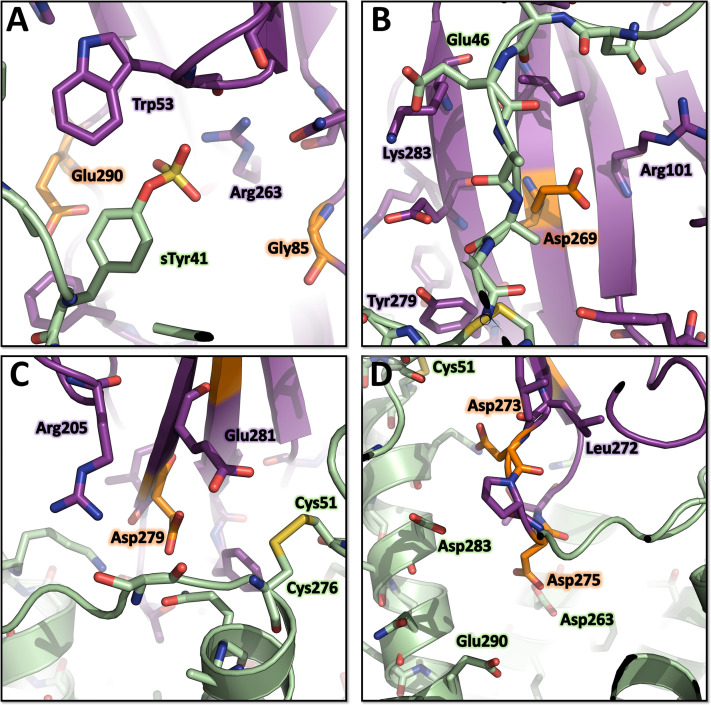
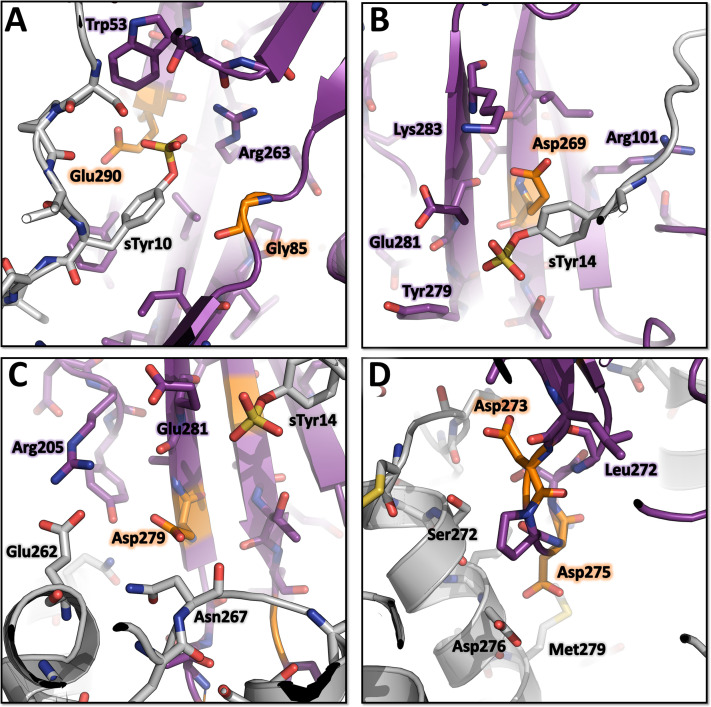
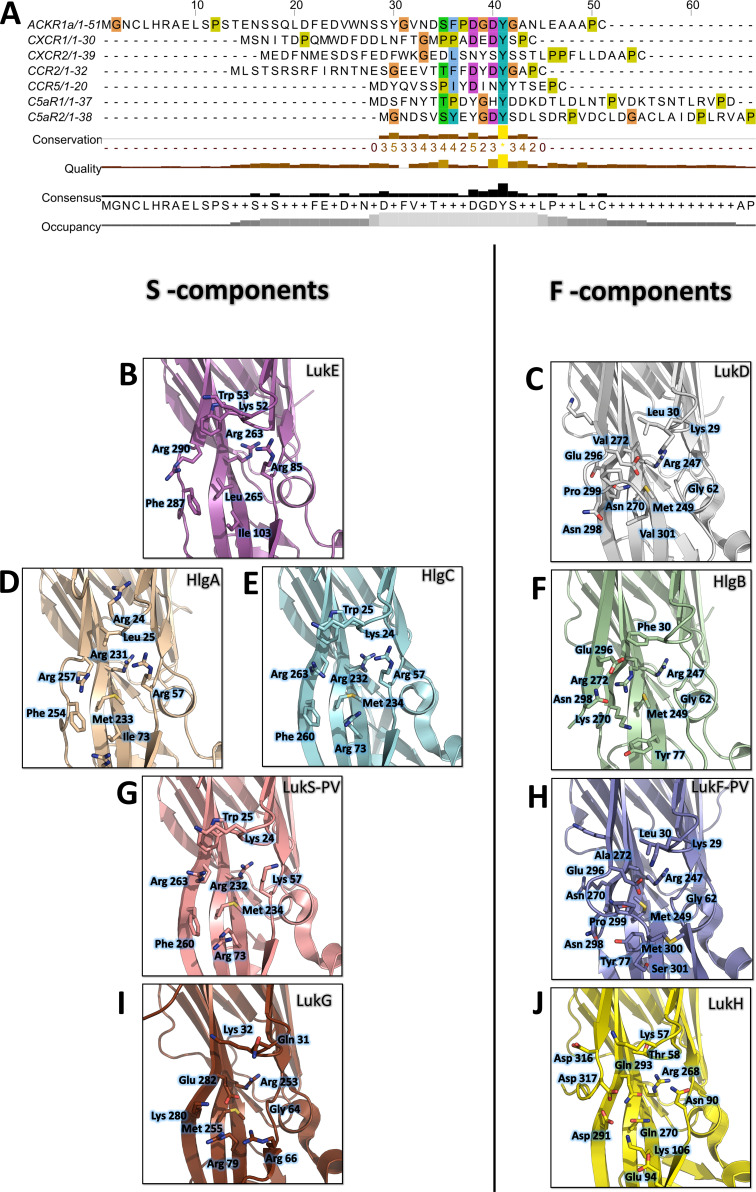
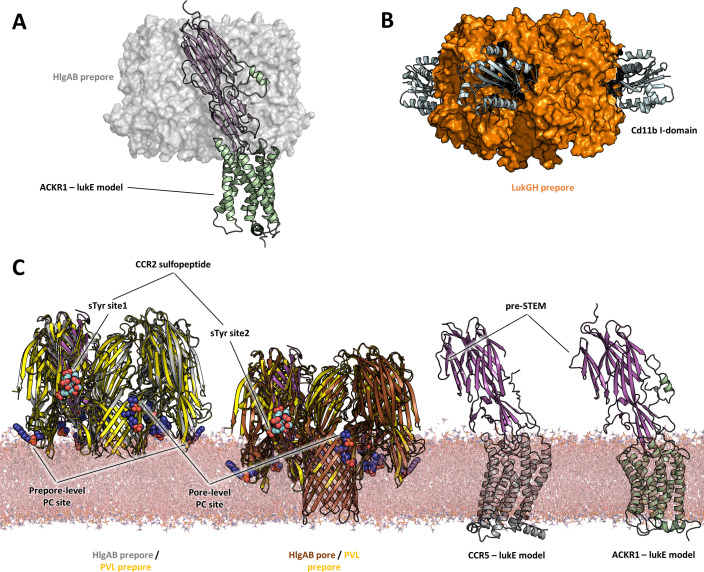
(SA) leukocidin ED (LukED) belongs to a family of bicomponent pore forming toxins that play important roles in SA immune evasion and nutrient acquisition. LukED targets specific G protein-coupled chemokine receptors to lyse human erythrocytes (red blood cells) and leukocytes (white blood cells). The first recognition step of receptors is critical for specific cell targeting and lysis. The structural and molecular bases for this mechanism are not well understood but could constitute essential information to guide antibiotic development. Here, we characterized the interaction of LukE with chemokine receptors ACKR1, CCR2, and CCR5 using a combination of structural, pharmacological, and computational approaches. First, crystal structures of LukE in complex with a small molecule mimicking sulfotyrosine side chain (p-cresyl sulfate) and with peptides containing sulfotyrosines issued from receptor sequences revealed the location of receptor sulfotyrosine binding sites in the toxins. Then, by combining previous and novel experimental data with protein docking, classical and accelerated weight histogram (AWH) molecular dynamics we propose models of the ACKR1-LukE and CCR5-LukE complexes. This work provides novel insights into chemokine receptor recognition by leukotoxins and suggests that the conserved sulfotyrosine binding pocket could be a target of choice for future drug development.
(金黄色葡萄球菌)白细胞毒素ED(LukED)属于双组分成孔毒素家族,在金黄色葡萄球菌的免疫逃逸和营养获取中发挥重要作用。LukED靶向特定的G蛋白偶联趋化因子受体,以裂解人类红细胞和白细胞。受体的首次识别步骤对于特定细胞靶向和裂解至关重要。这种机制的结构和分子基础尚不清楚,但可能构成指导抗生素开发的重要信息。在这里,我们结合结构、药理学和计算方法,对LukE与趋化因子受体ACKR1、CCR2和CCR5的相互作用进行了表征。首先,LukE与模拟磺基酪氨酸侧链的小分子(对甲酚硫酸盐)以及含有受体序列中磺基酪氨酸的肽形成的晶体结构揭示了毒素中受体磺基酪氨酸结合位点的位置。然后,通过将先前和新的实验数据与蛋白质对接、经典和加速权重直方图(AWH)分子动力学相结合,我们提出了ACKR1-LukE和CCR5-LukE复合物的模型。这项工作为白细胞毒素对趋化因子受体的识别提供了新的见解,并表明保守的磺基酪氨酸结合口袋可能是未来药物开发的首选靶点。