Department of Biological Sciences, Vanderbilt University, Nashville, United States.
Department of Cell and Developmental Biology, Vanderbilt University, Nashville, United States.
Elife. 2022 Apr 22;11:e71634. doi: 10.7554/eLife.71634.
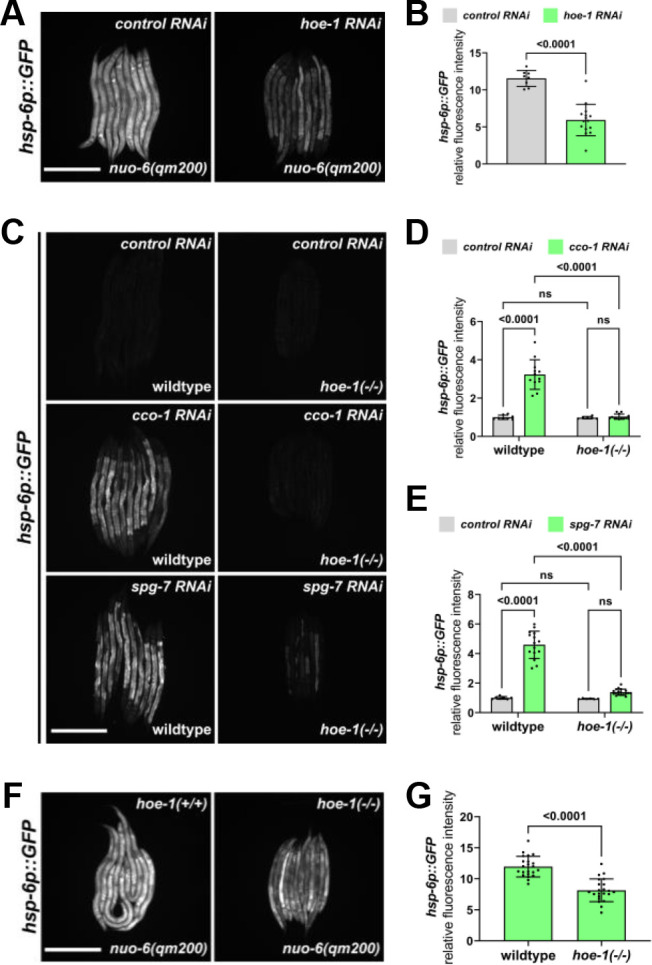
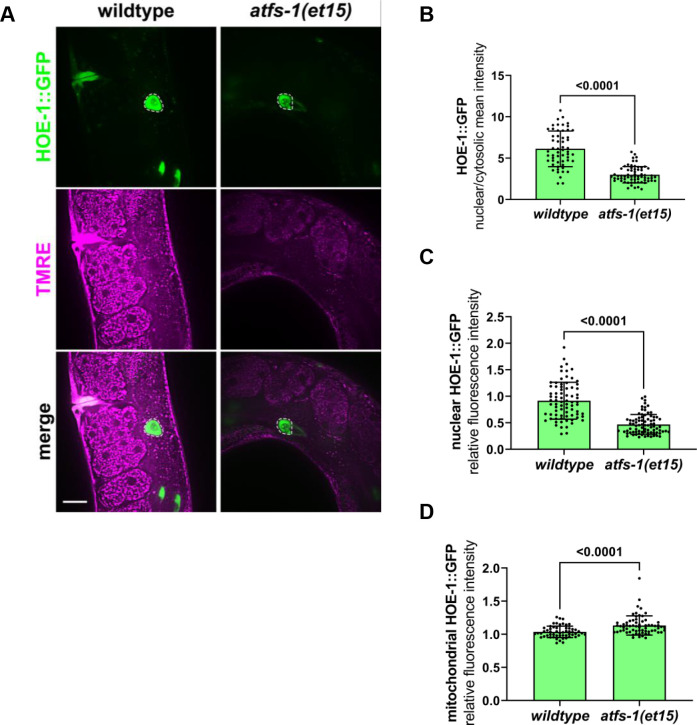
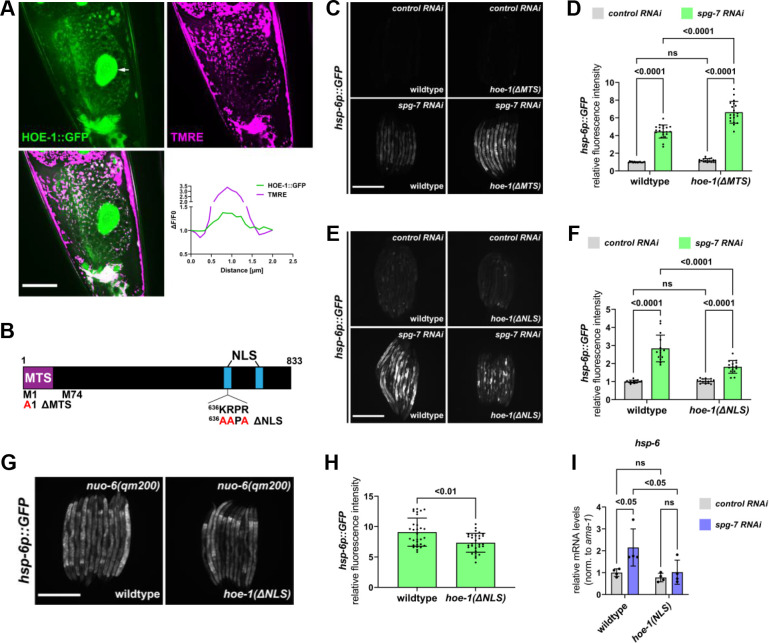
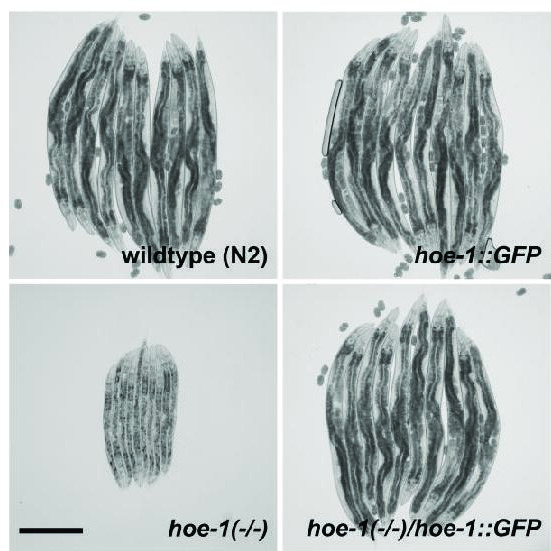
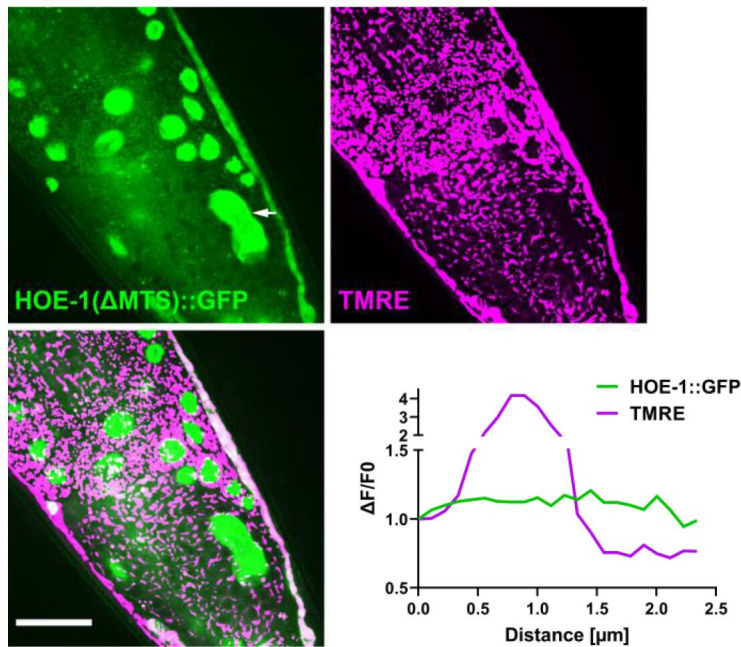
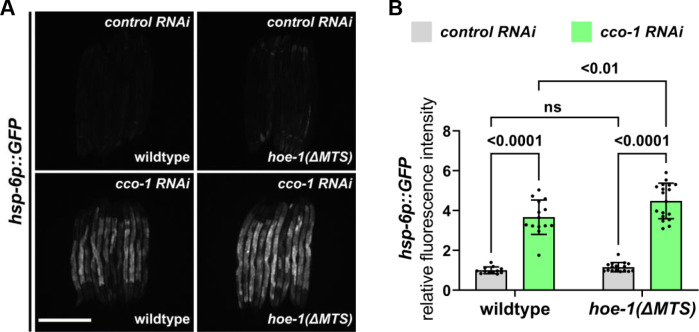
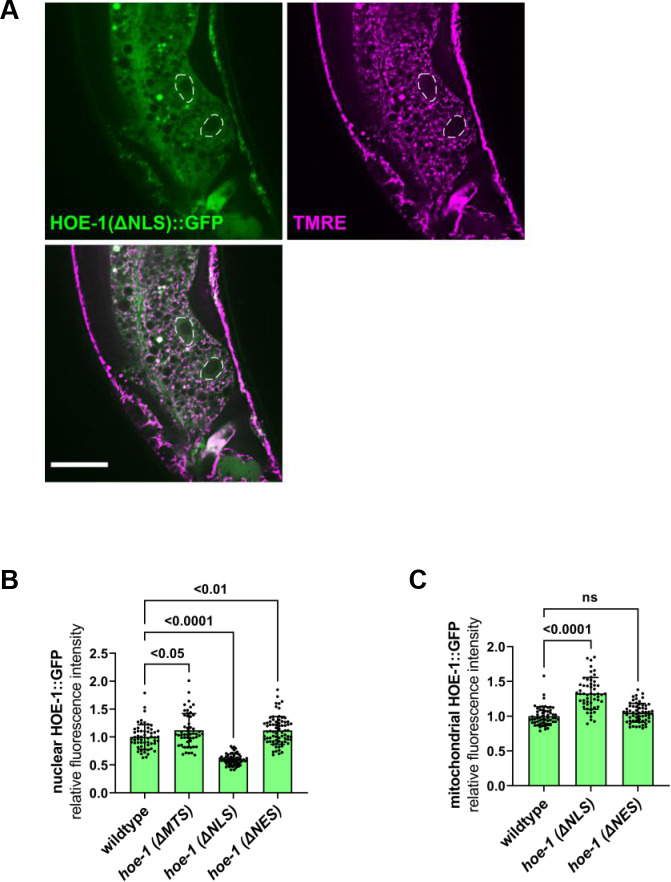
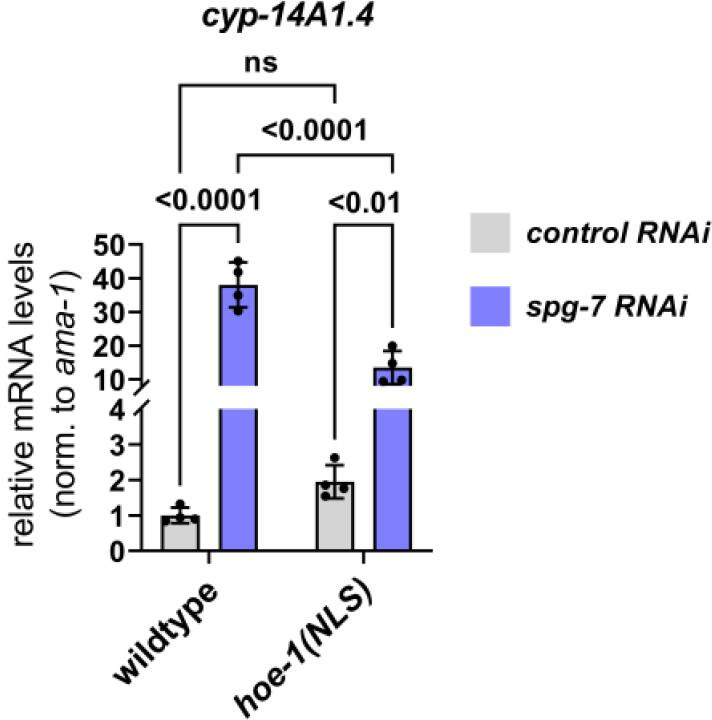
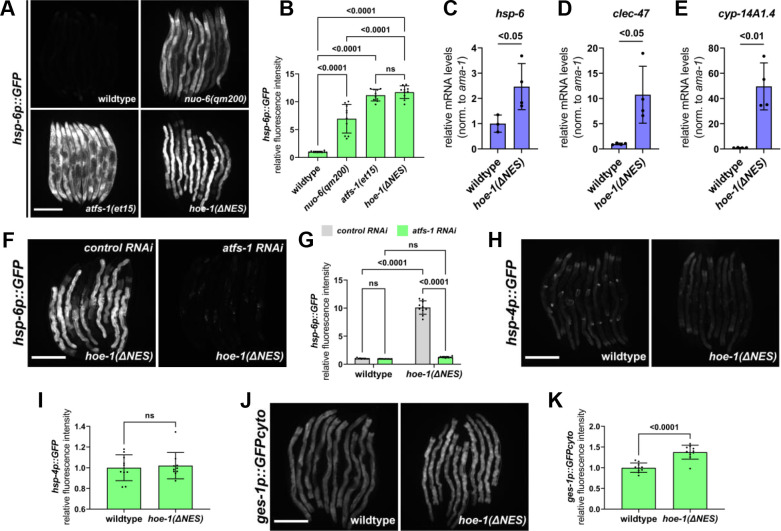
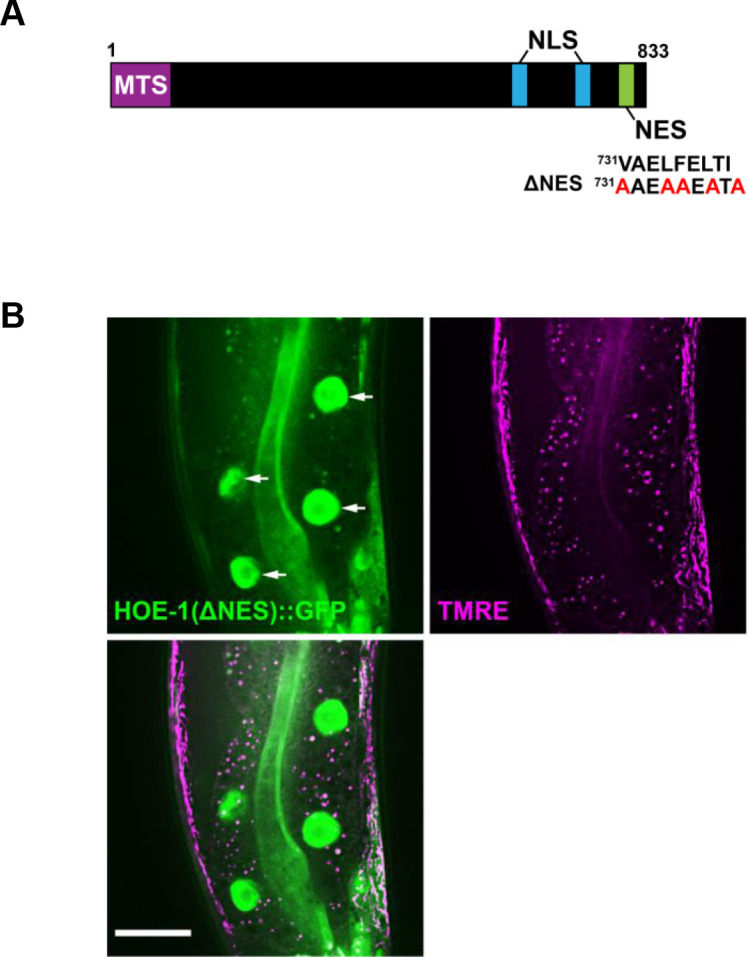
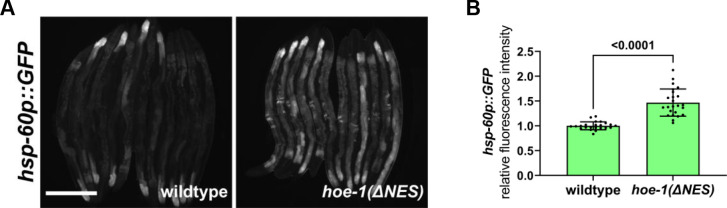
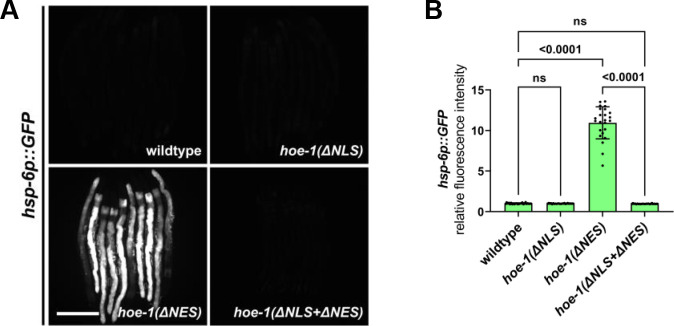
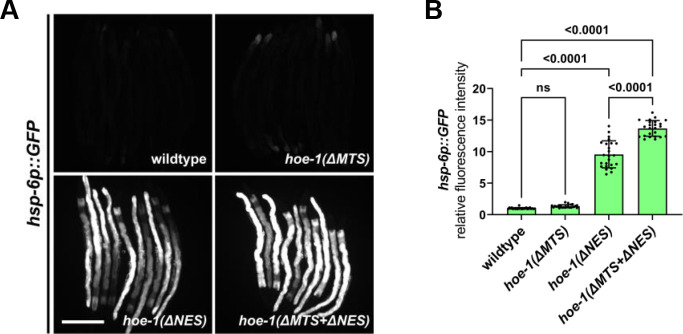
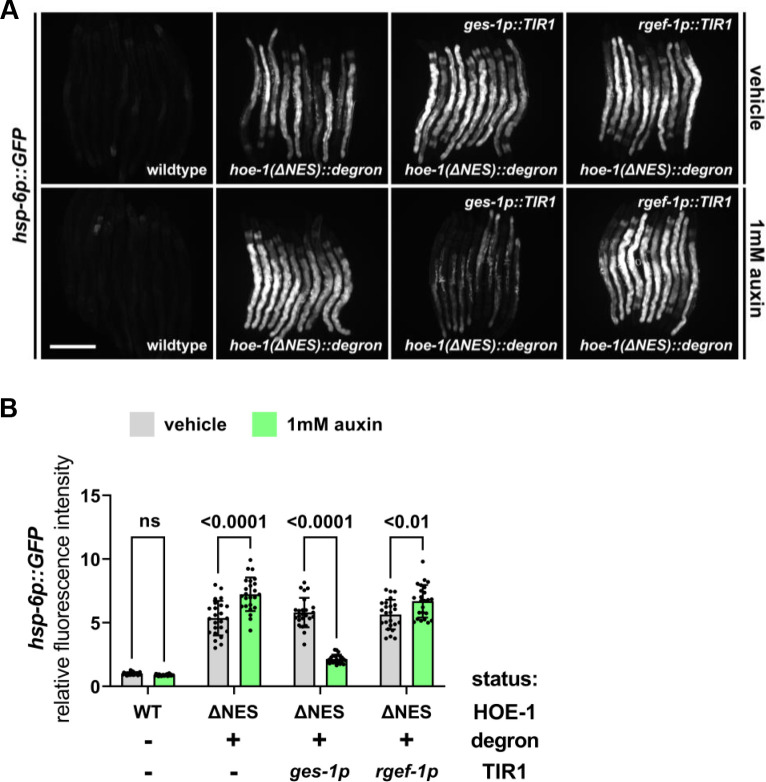
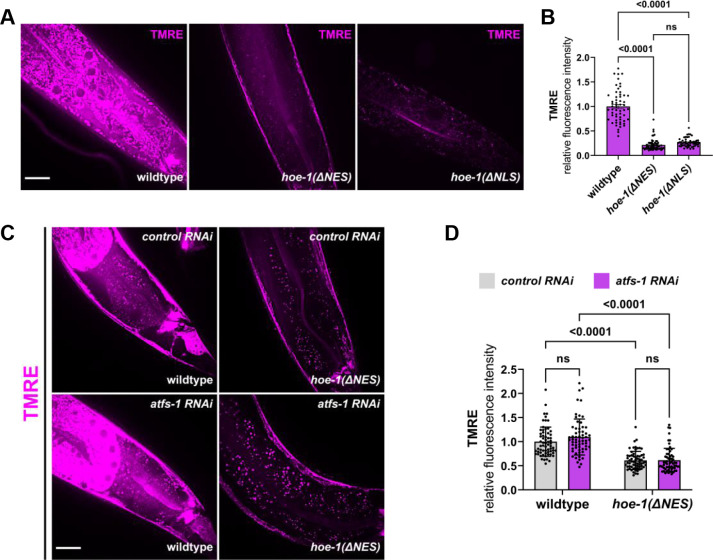
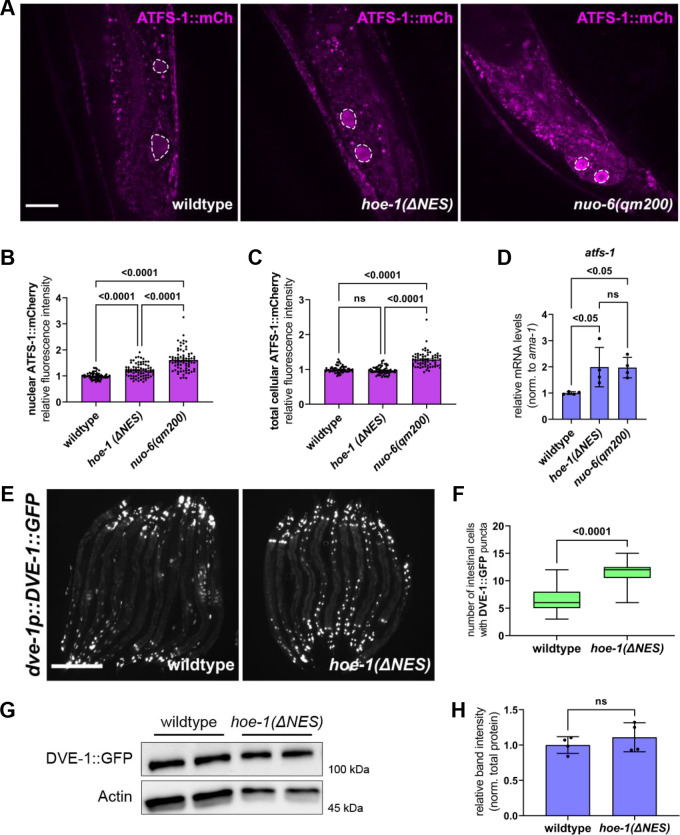
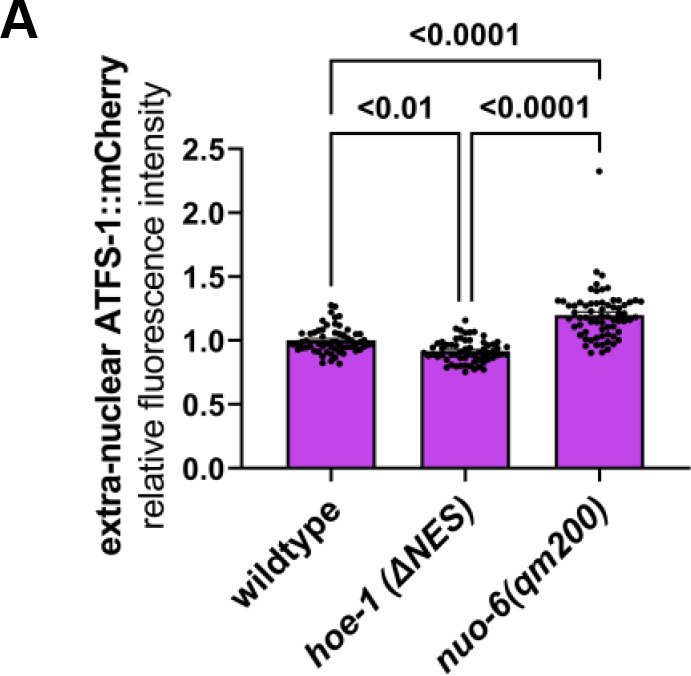
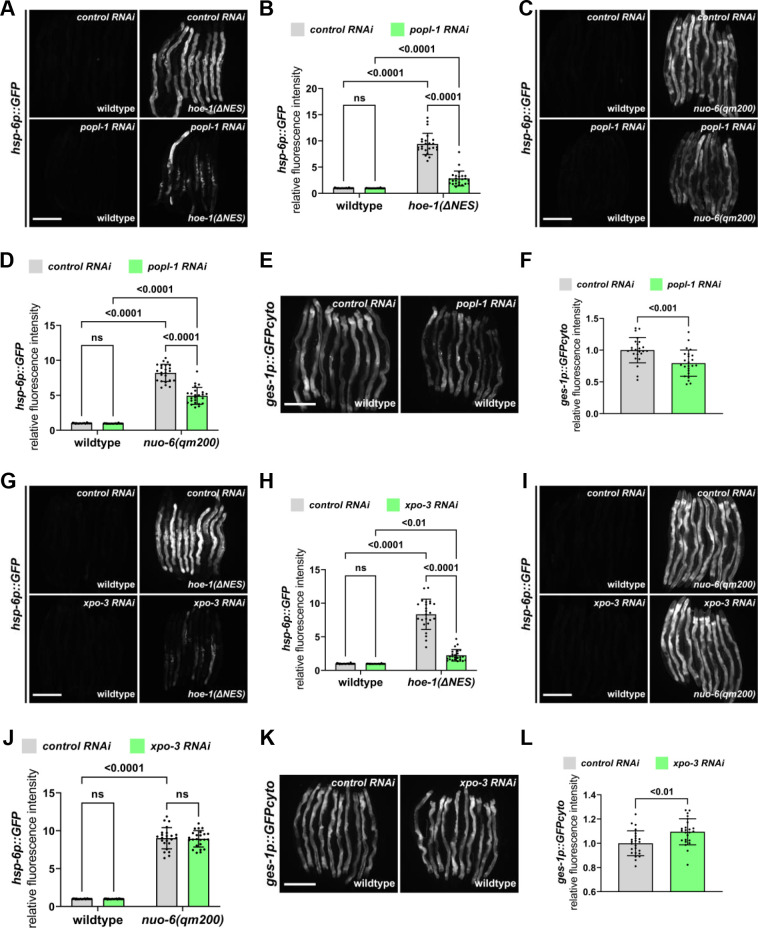
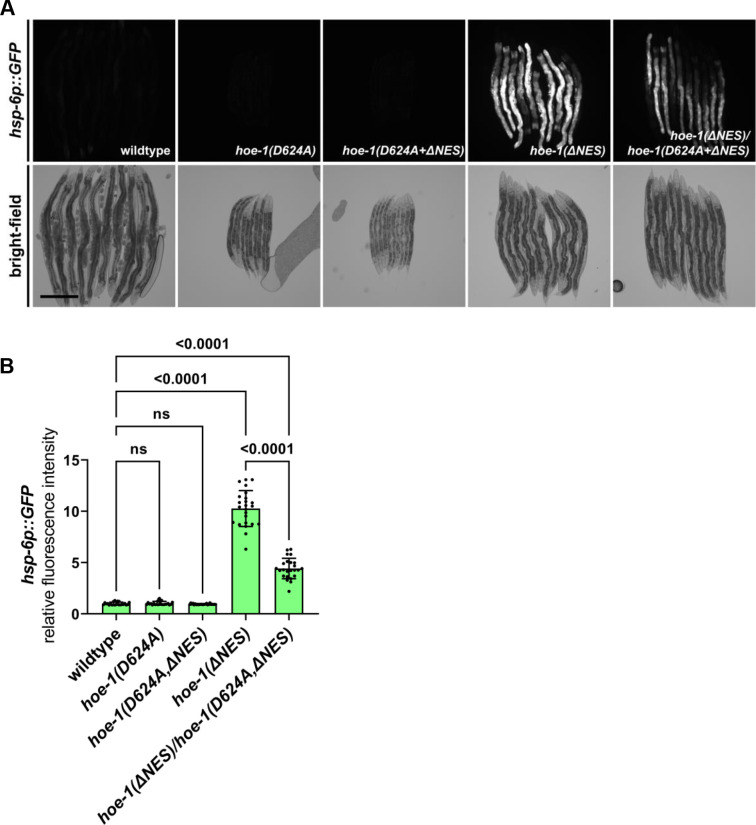
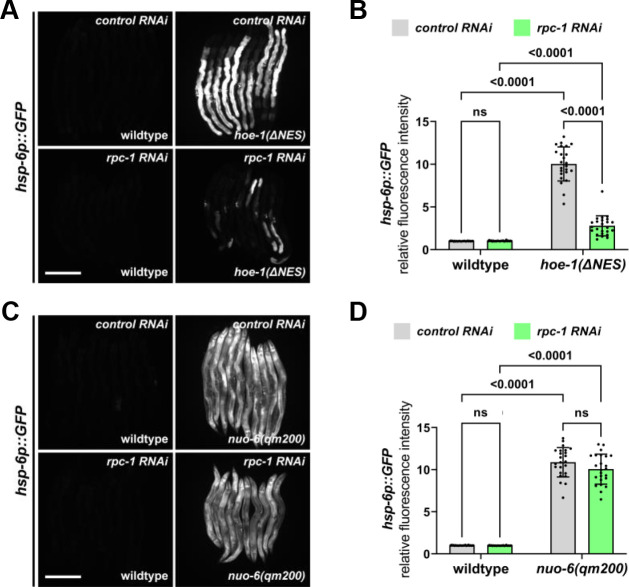
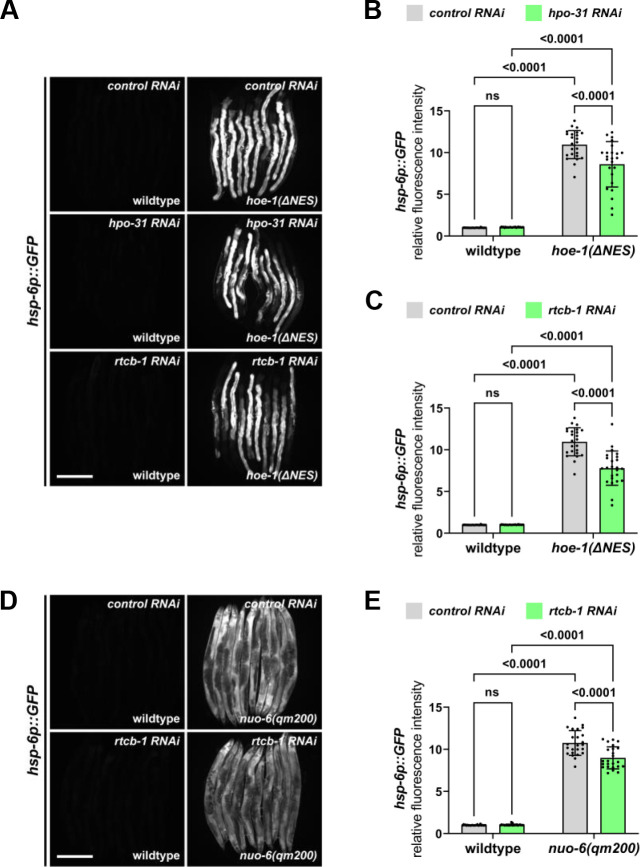
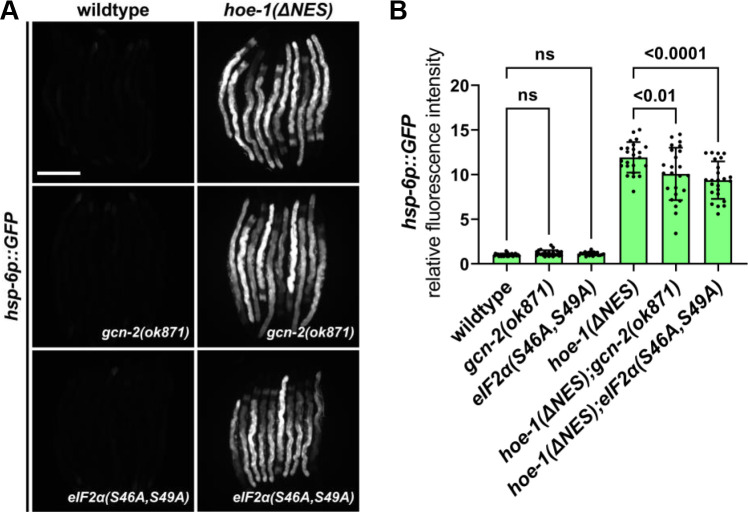
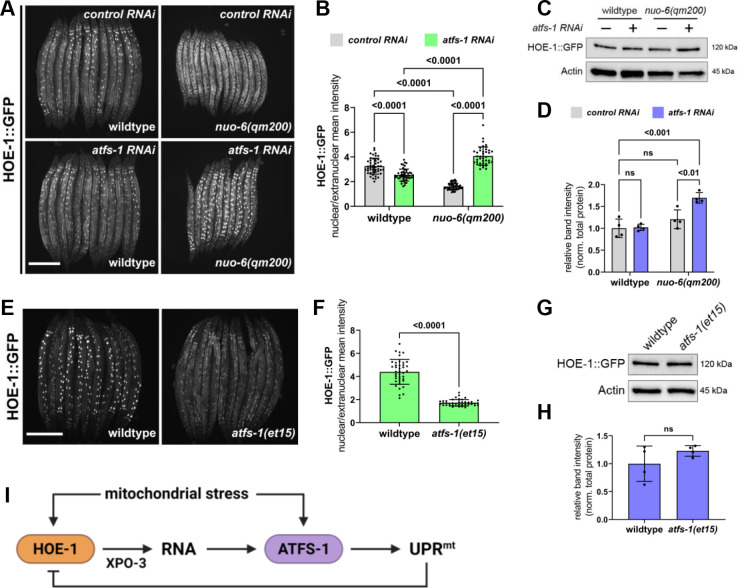
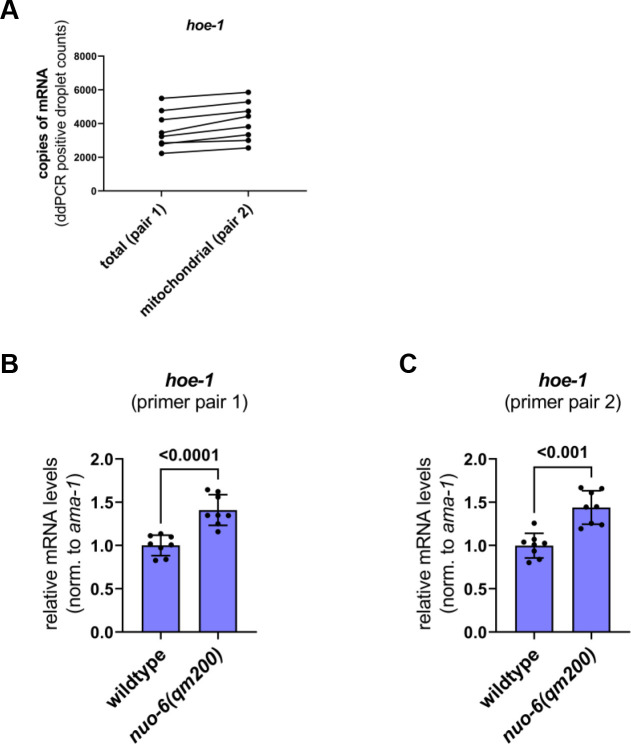
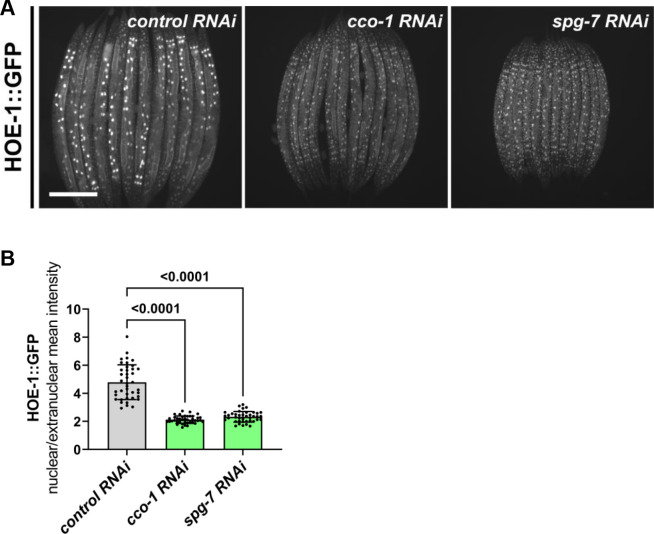
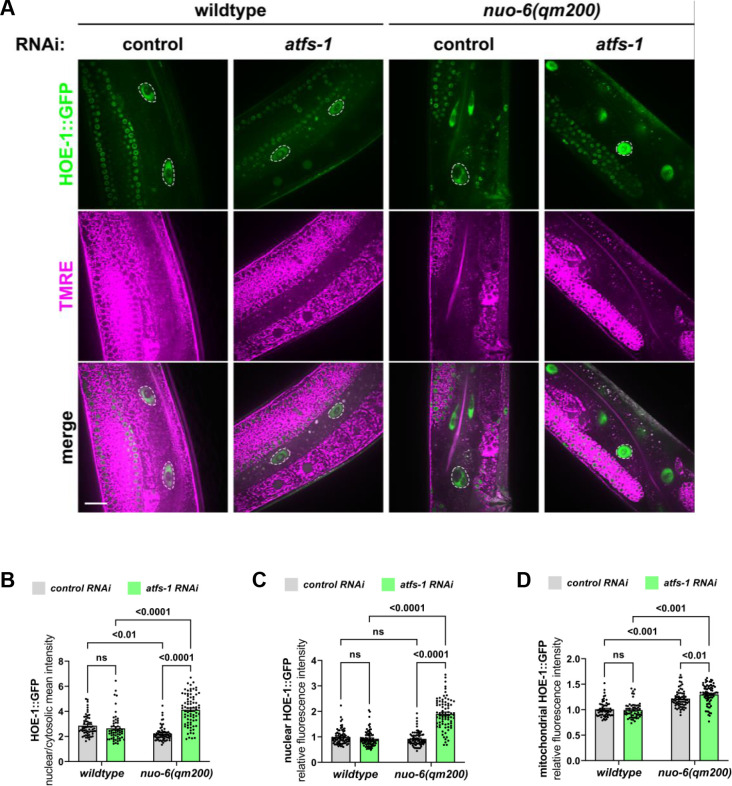
The mitochondrial unfolded protein response (UPR) has emerged as a predominant mechanism that preserves mitochondrial function. Consequently, multiple pathways likely exist to modulate UPR. We discovered that the tRNA processing enzyme, homolog of ELAC2 (HOE-1), is key to UPR regulation in . We find that nuclear HOE-1 is necessary and sufficient to robustly activate UPR. We show that HOE-1 acts via transcription factors ATFS-1 and DVE-1 that are crucial for UPR. Mechanistically, we show that HOE-1 likely mediates its effects via tRNAs, as blocking tRNA export prevents HOE-1-induced UPR. Interestingly, we find that HOE-1 does not act via the integrated stress response, which can be activated by uncharged tRNAs, pointing toward its reliance on a new mechanism. Finally, we show that the subcellular localization of HOE-1 is responsive to mitochondrial stress and is subject to negative regulation via ATFS-1. Together, we have discovered a novel RNA-based cellular pathway that modulates UPR.
线粒体未折叠蛋白反应 (UPR) 已成为维持线粒体功能的主要机制。因此,可能存在多种途径来调节 UPR。我们发现 tRNA 加工酶同源物 ELAC2 (HOE-1) 是 中 UPR 调节的关键。我们发现核 HOE-1 是激活 UPR 所必需且充分的。我们表明 HOE-1 通过 ATFS-1 和 DVE-1 转录因子发挥作用,这些因子对于 UPR 至关重要。在机制上,我们表明 HOE-1 可能通过 tRNA 介导其作用,因为阻止 tRNA 输出可防止 HOE-1 诱导的 UPR。有趣的是,我们发现 HOE-1 不通过整合应激反应起作用,整合应激反应可被未结合的 tRNA 激活,这表明它依赖于一种新的机制。最后,我们表明 HOE-1 的亚细胞定位对线粒体应激有反应,并通过 ATFS-1 受到负调节。总之,我们发现了一种新的基于 RNA 的细胞途径,可调节 UPR。