Department of Biochemistry and Molecular Biology, Johns Hopkins University Bloomberg School of Public Health, Baltimore, Maryland 21205, United States.
Department of Physics, Wake Forest University, Winston-Salem, North Carolina 27109, United States.
Biochemistry. 2022 May 17;61(10):833-842. doi: 10.1021/acs.biochem.2c00096. Epub 2022 Apr 28.
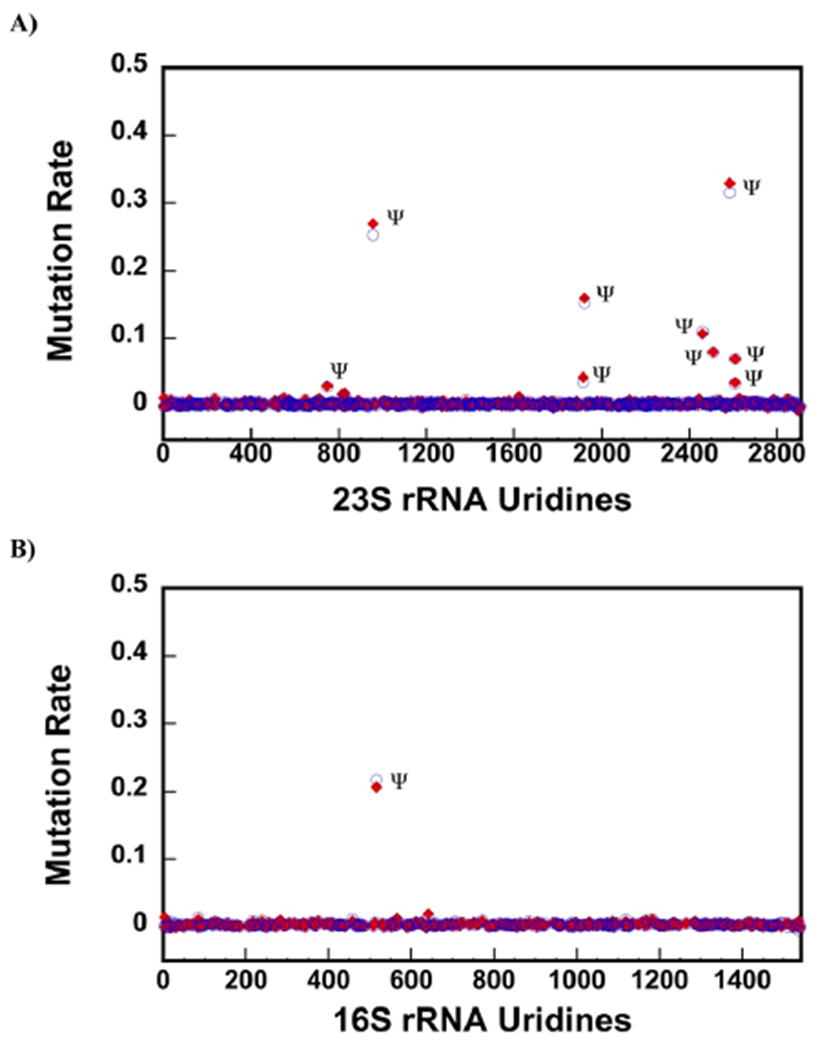
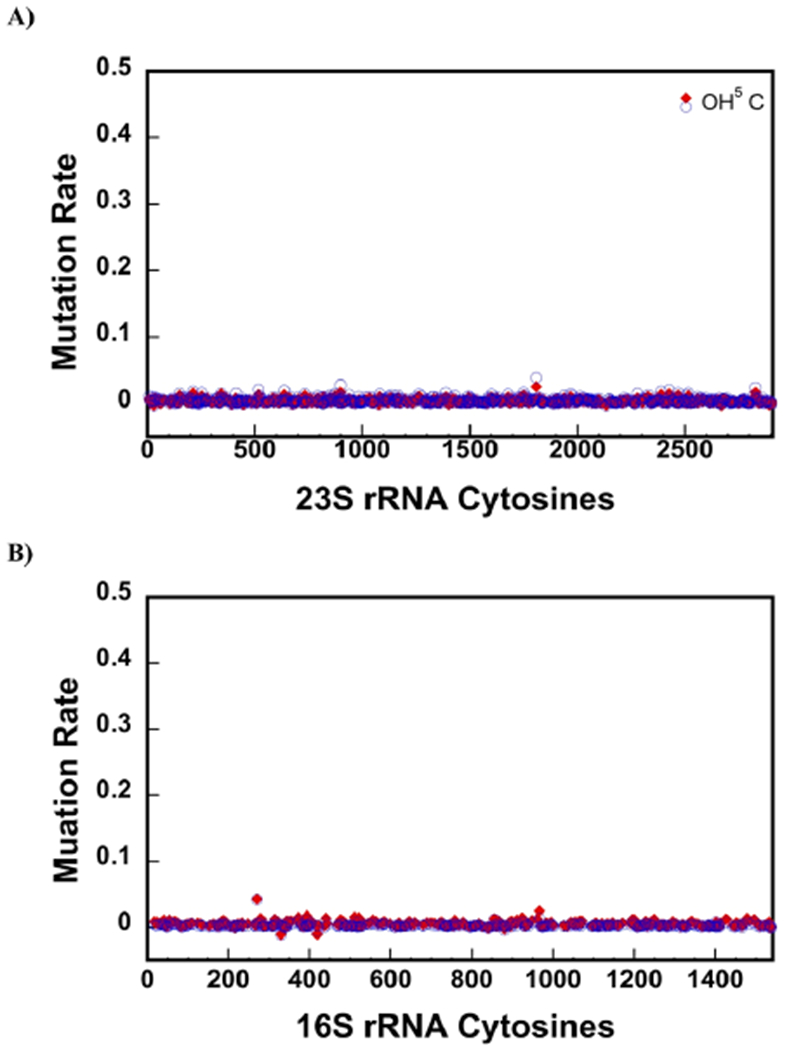
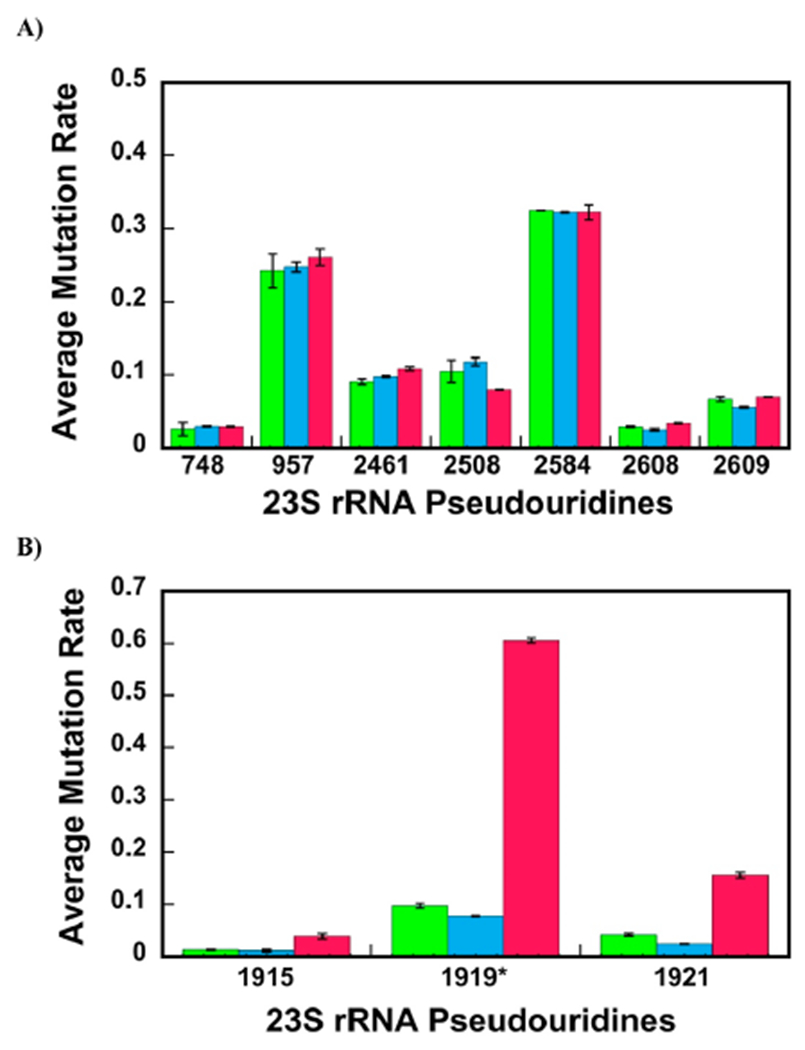
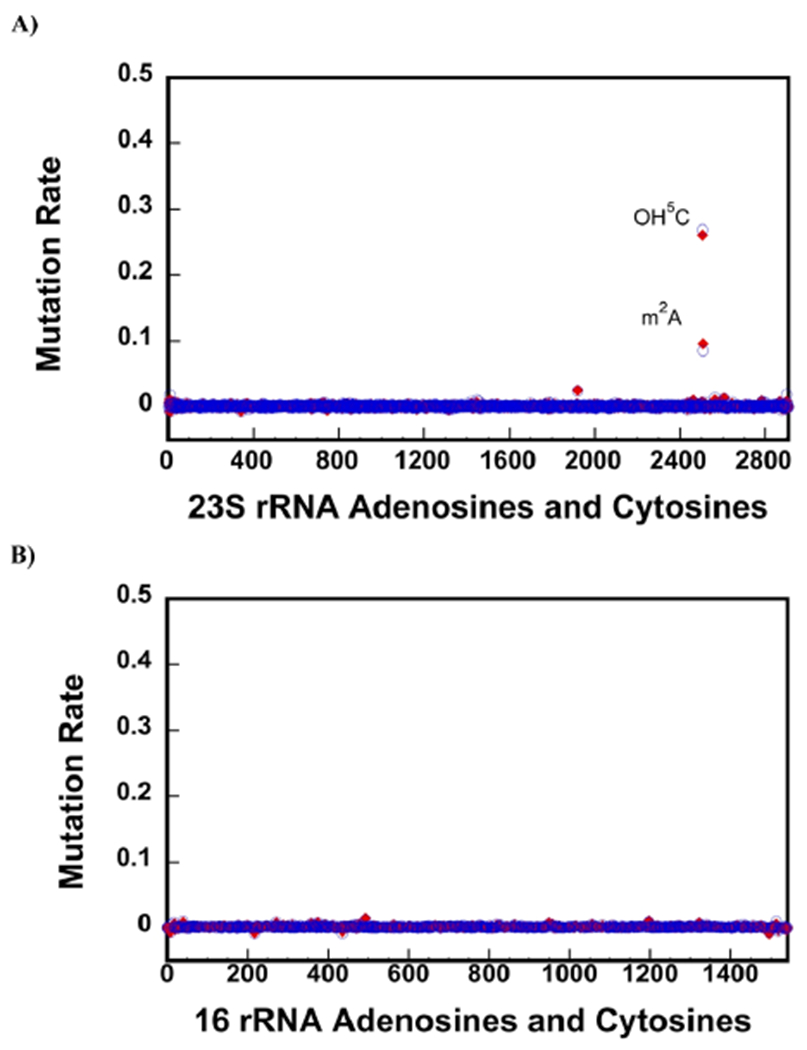
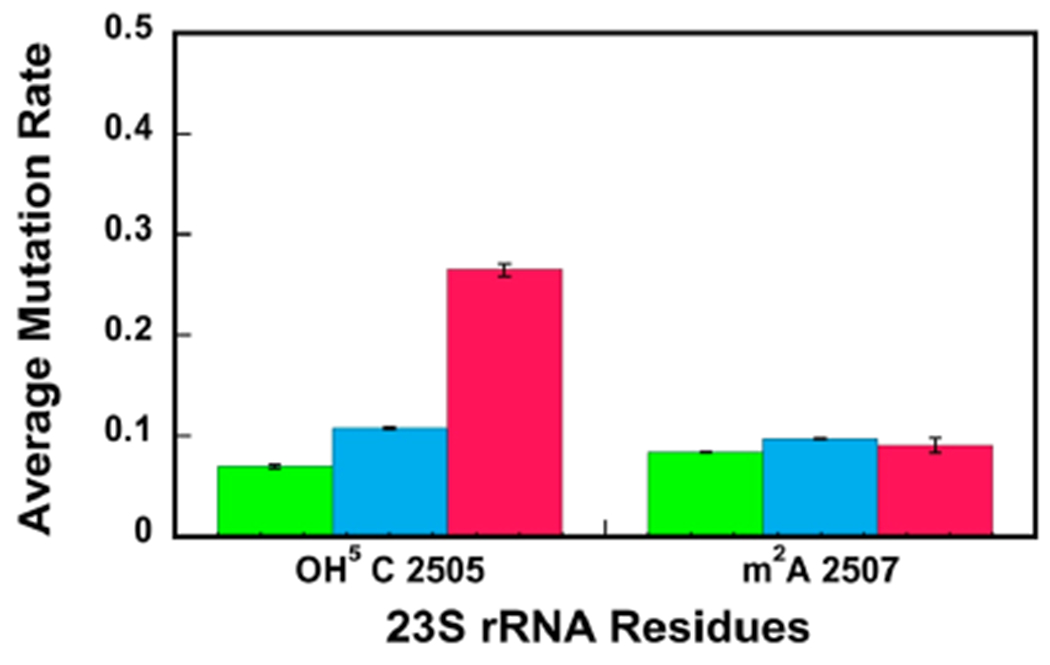
23S ribosomal RNA (rRNA) of 50S large ribosome subunit contains 26 post-transcriptionally modified nucleosides. Here, we determine the extent of modifications in the 35S and 45S large subunit intermediates, accumulating in cells expressing the helicase inactive DbpA protein, R331A, and the native 50S large subunit. The modifications we characterized are 3-methylpseudouridine, 2-methyladenine, 5-hydroxycytidine, and nine pseudouridines. These modifications were detected using 1-cyclohexyl-3-(2-morpholinoethyl)carbodiimide metho--toluenesulfonate (CMCT) treatment followed by alkaline treatment. In addition, KMnO treatment of 23S rRNA was employed to detect 5-hydroxycytidine modification. CMCT and KMnO treatments produce chemical changes in modified nucleotides that cause reverse transcriptase misincorporations and deletions, which were detected employing next-generation sequencing. Our results show that the 2-methyladenine modification and seven uridines to pseudouridine isomerizations are present in both the 35S and 45S to similar extents as in the 50S. Hence, the enzymes that perform these modifications, namely, RluA, RluB, RluC, RluE, RluF, and RlmN, have already acted in the intermediates. Two uridines to pseudouridine isomerizations, the 3-methylpseudouridine and 5-hydroxycytidine modifications, are significantly less present in the 35S and 45S, as compared to the 50S. Therefore, the enzymes that incorporate these modifications, RluD, RlmH, and RlhA, are in the process of modifying the 35S and 45S or will incorporate these modifications during the later stages of ribosome assembly. Our study employs a novel high throughput and single nucleotide resolution technique for the detection of 2-methyladenine and two novel high throughput and single nucleotide resolution techniques for the detection of 5-hydroxycytidine.
23S 核糖体 RNA(rRNA)的 50S 大亚基包含 26 个转录后修饰核苷。在这里,我们确定了在表达无解旋酶活性的 DbpA 蛋白 R331A 和天然 50S 大亚基的细胞中积累的 35S 和 45S 大亚基中间产物的修饰程度。我们鉴定的修饰包括 3-甲基假尿嘧啶、2-甲基腺嘌呤、5-羟化胞嘧啶和 9 个假尿嘧啶。这些修饰是通过 1-环己基-3-(2-吗啉代乙基)碳二亚胺甲--对甲苯磺酸酯 (CMCT)处理后用碱性处理来检测的。此外,采用 KMnO 处理 23S rRNA 来检测 5-羟化胞嘧啶修饰。CMCT 和 KMnO 处理会导致修饰核苷酸发生化学变化,从而导致逆转录酶错误掺入和缺失,这可以通过下一代测序来检测。我们的结果表明,2-甲基腺嘌呤修饰和 7 个尿嘧啶到假尿嘧啶的异构化在 35S 和 45S 中与 50S 中存在的程度相似。因此,执行这些修饰的酶,即 RluA、RluB、RluC、RluE、RluF 和 RlmN,已经在中间体中发挥了作用。与 50S 相比,35S 和 45S 中两个尿嘧啶到假尿嘧啶的异构化、3-甲基假尿嘧啶和 5-羟化胞嘧啶修饰的程度明显较低。因此,掺入这些修饰的酶,即 RluD、RlmH 和 RlhA,正在修饰 35S 和 45S,或者将在核糖体组装的后期阶段掺入这些修饰。我们的研究采用了一种新的高通量和单核苷酸分辨率技术来检测 2-甲基腺嘌呤,并采用两种新的高通量和单核苷酸分辨率技术来检测 5-羟化胞嘧啶。