Center for Integrated Protein Research at the Department of Chemistry, Technische Universität München, Lichtenbergstr. 4, 85748, Garching, Germany.
Coriolis Pharma Research GmbH, Fraunhoferstraße 18b, 82152, Martinsried, Germany.
Sci Rep. 2022 May 28;12(1):8984. doi: 10.1038/s41598-022-12736-x.
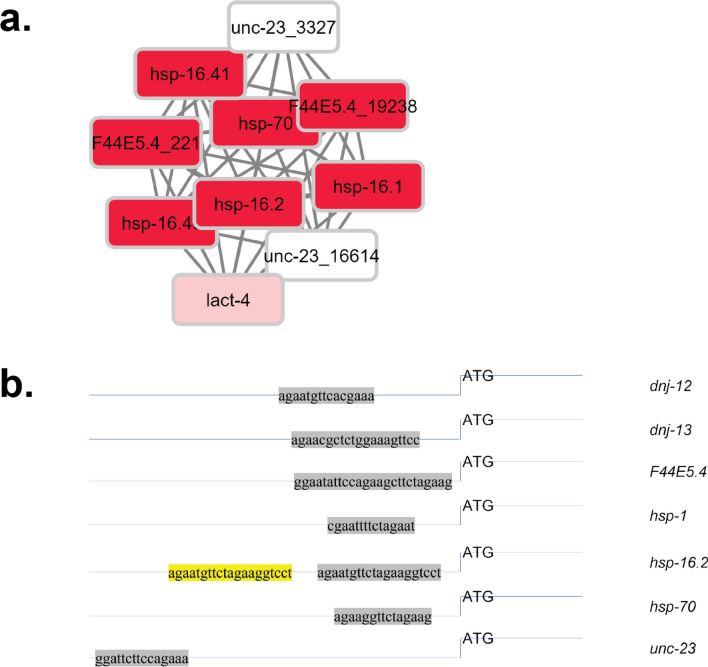
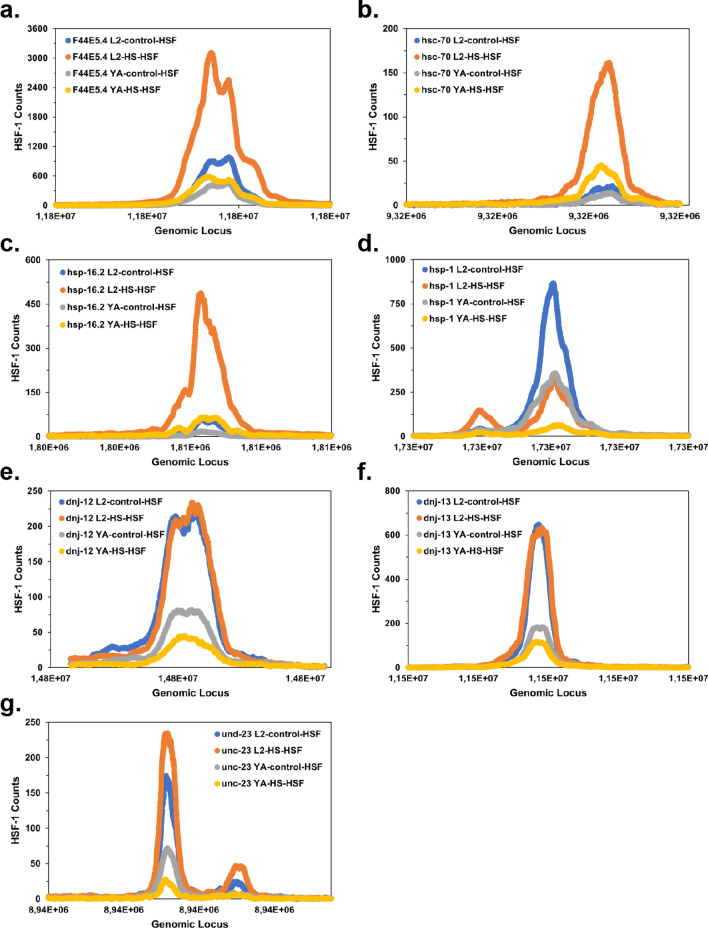
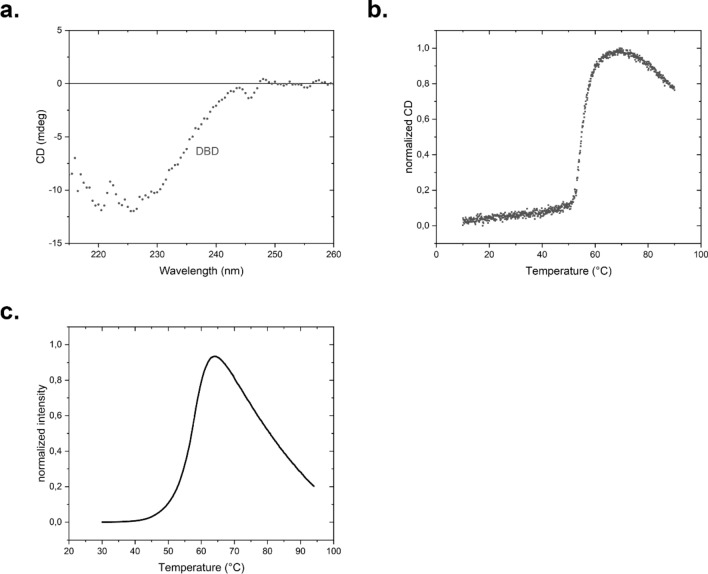
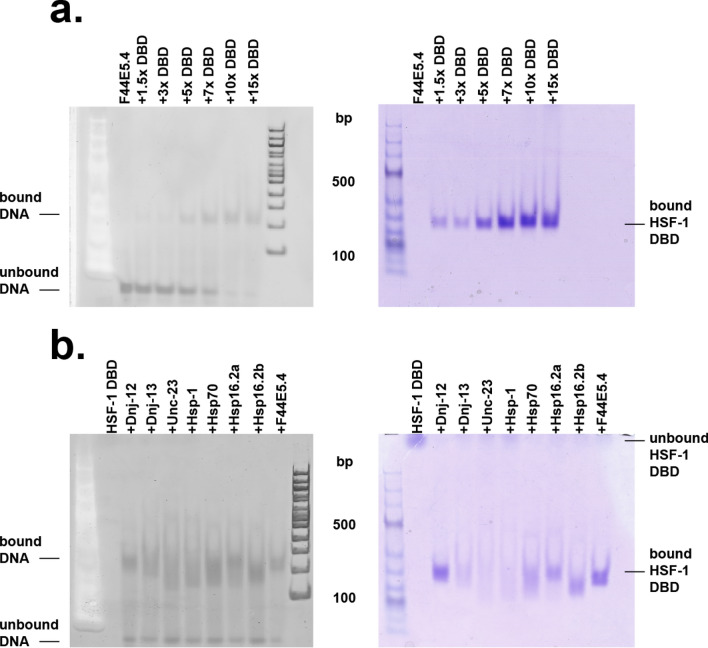
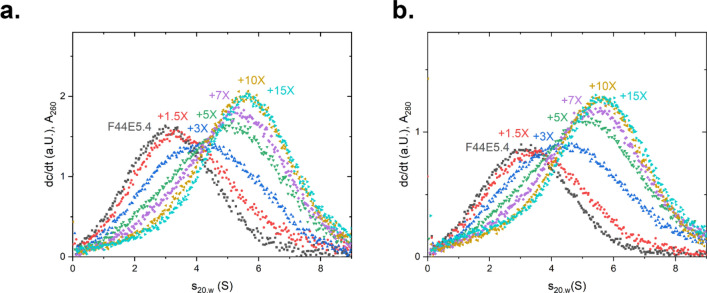
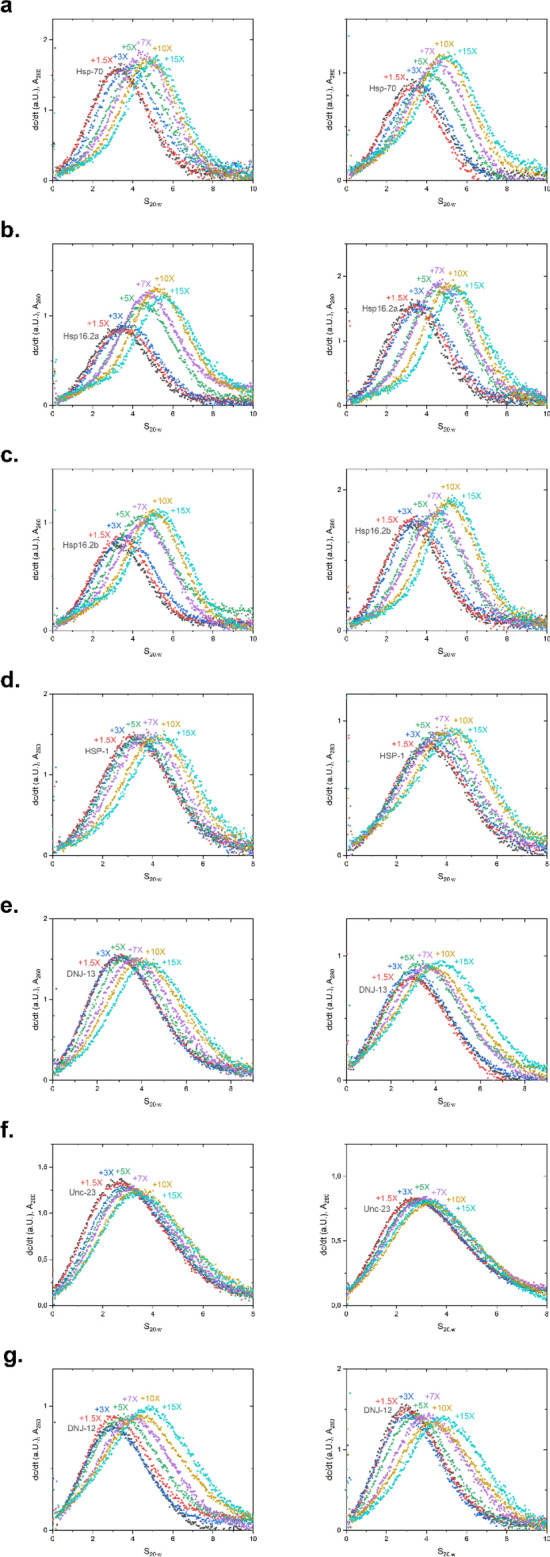
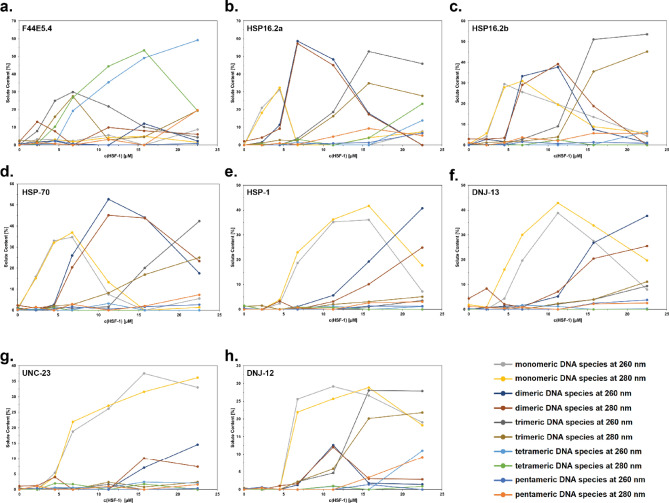
The protein HSF-1 is the controlling transcription factor of the heat-shock response (HSR). Its binding to the heat-shock elements (HSEs) induces the strong upregulation of conserved heat-shock proteins, including Hsp70s, Hsp40s and small HSPs. Next to these commonly known HSPs, more than 4000 other HSEs are found in the promoter regions of C. elegans genes. In microarray experiments, few of the HSE-containing genes are specifically upregulated during the heat-shock response. Most of the 4000 HSE-containing genes instead are unaffected by elevated temperatures and coexpress with genes unrelated to the HSR. This is also the case for several genes related to the HSP chaperone system, like dnj-12, dnj-13, and hsp-1. Interestingly, several promoters of the dedicated HSR-genes, like F44E5.4p, hsp-16.48p or hsp-16.2p, contain extended HSEs in their promoter region, composed of four or five HSE-elements instead of the common trimeric HSEs. We here aim at understanding how HSF-1 interacts with the different promoter regions. To this end we purify the nematode HSF-1 DBD and investigate the interaction with DNA sequences containing these regions. EMSA assays suggest that the HSF-1 DBD interacts with most of these HSE-containing dsDNAs, but with different characteristics. We employ sedimentation analytical ultracentrifugation (SV-AUC) to determine stoichiometry, affinity, and cooperativity of HSF-1 DBD binding to these HSEs. Interestingly, most HSEs show cooperative binding of the HSF-1 DBD with up to five DBDs being bound. In most cases binding to the HSEs of inducible promoters is stronger, even though the consensus scores are not always higher. The observed high affinity of HSF-1 DBD to the non-inducible HSEs of dnj-12, suggests that constitutive expression may be supported from some promoter regions, a fact that is evident for this transcription factor, that is essential also under non-stress conditions.
HSF-1 蛋白是热休克反应 (HSR) 的调控转录因子。它与热休克元件 (HSEs) 结合,诱导保守的热休克蛋白(包括 Hsp70s、Hsp40s 和小分子 HSPs)的强烈上调。除了这些常见的 HSPs 之外,在 C. elegans 基因的启动子区域还发现了超过 4000 个其他的 HSEs。在微阵列实验中,在热休克反应期间,只有少数含有 HSE 的基因被特异性地上调。相反,大多数含有 4000 个 HSE 的基因不受高温的影响,并与与 HSR 无关的基因共同表达。与 HSP 伴侣系统相关的几个基因也是如此,如 dnj-12、dnj-13 和 hsp-1。有趣的是,几个专门的 HSR 基因的启动子,如 F44E5.4p、hsp-16.48p 或 hsp-16.2p,在其启动子区域含有扩展的 HSE,由四个或五个 HSE 元件组成,而不是常见的三聚体 HSE。我们在这里旨在了解 HSF-1 如何与不同的启动子区域相互作用。为此,我们纯化了线虫 HSF-1 DBD,并研究了与含有这些区域的 DNA 序列的相互作用。EMSA 实验表明,HSF-1 DBD 与大多数含有这些 HSE 的 dsDNA 相互作用,但具有不同的特征。我们采用沉降分析超速离心(SV-AUC)来确定 HSF-1 DBD 与这些 HSE 结合的化学计量、亲和力和协同性。有趣的是,大多数 HSE 表现出 HSF-1 DBD 的协同结合,多达五个 DBD 被结合。在大多数情况下,诱导型启动子的 HSE 结合更强,尽管共识评分并不总是更高。DNJ-12 的非诱导性 HSE 与 HSF-1 DBD 的高亲和力表明,组成型表达可能得到一些启动子区域的支持,这对于这个转录因子来说是一个明显的事实,它在非应激条件下也是必不可少的。