Department of Biological Sciences, Vanderbilt University, Nashville, United States.
Department of Cell and Developmental Biology, Vanderbilt University, Nashville, United States.
Elife. 2022 Jul 29;11:e80911. doi: 10.7554/eLife.80911.
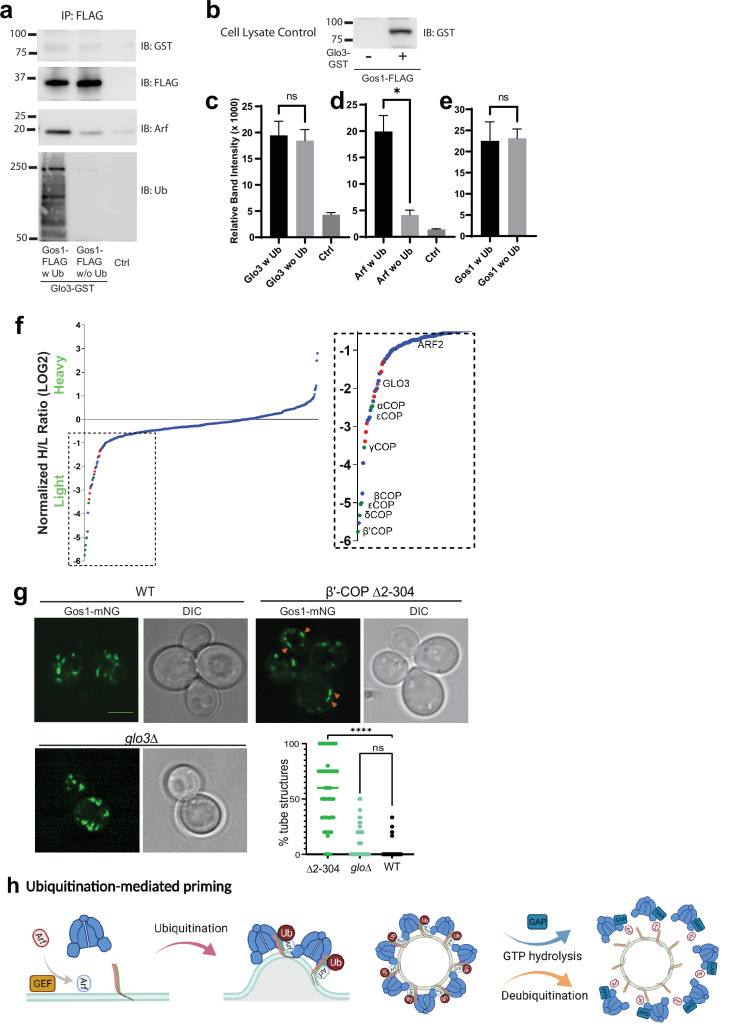
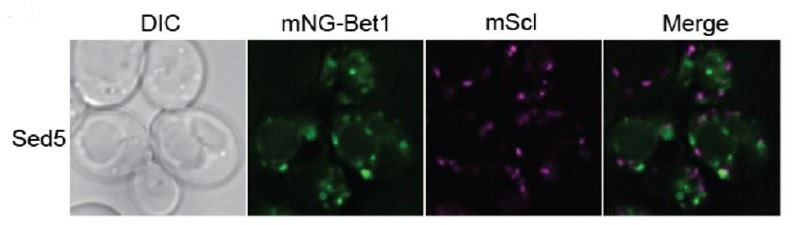
Deciphering mechanisms controlling SNARE localization within the Golgi complex is crucial to understanding protein trafficking patterns within the secretory pathway. SNAREs are also thought to prime coatomer protein I (COPI) assembly to ensure incorporation of these essential cargoes into vesicles, but the regulation of these events is poorly understood. Here, we report roles for ubiquitin recognition by COPI in SNARE trafficking and in stabilizing interactions between Arf, COPI, and Golgi SNAREs in . The ability of COPI to bind ubiquitin, but not the dilysine motif, through its N-terminal WD repeat domain of β'-COP or through an unrelated ubiquitin-binding domain is essential for the proper localization of Golgi SNAREs Bet1 and Gos1. We find that COPI, the ArfGAP Glo3, and multiple Golgi SNAREs are ubiquitinated. Notably, the binding of Arf and COPI to Gos1 is markedly enhanced by ubiquitination of these components. Glo3 is proposed to prime COPI-SNARE interactions; however, Glo3 is not enriched in the ubiquitin-stabilized SNARE-Arf-COPI complex but is instead enriched with COPI complexes that lack SNAREs. These results support a new model for how posttranslational modifications drive COPI priming events crucial for Golgi SNARE localization.
解析控制 SNARE 在高尔基体复合物中定位的机制对于理解分泌途径中蛋白质运输模式至关重要。SNARE 还被认为可以启动衣被蛋白 I(COPI)组装,以确保这些必需货物被纳入囊泡中,但这些事件的调节机制尚不清楚。在这里,我们报告了 COPI 通过其 β'-COP 的 N 端 WD 重复结构域或通过不相关的泛素结合结构域识别泛素在 SNARE 运输和稳定 Arf、COPI 和高尔基体 SNARE 之间相互作用中的作用。COPI 能够结合泛素,但不能结合二赖氨酸基序,这对于高尔基体 SNAREs Bet1 和 Gos1 的正确定位是必不可少的。我们发现 COPI、ArfGAP Glo3 和多个高尔基体 SNARE 都被泛素化了。值得注意的是,这些成分的泛素化显著增强了 Arf 和 COPI 与 Gos1 的结合。Glo3 被提议为 COPI-SNARE 相互作用的启动子;然而,Glo3 没有在稳定的 SNARE-Arf-COPI 复合物中富集,而是在缺乏 SNARE 的 COPI 复合物中富集。这些结果支持了一个新的模型,即翻译后修饰如何驱动 COPI 引发对于高尔基体 SNARE 定位至关重要的事件。