Department of Biology/Chemistry, Structural Biology section, Osnabrück University, Osnabrück, Germany.
Department of Biology/Chemistry, Biochemistry section, Osnabrück University, Osnabrück, Germany.
Elife. 2022 Sep 13;11:e80901. doi: 10.7554/eLife.80901.
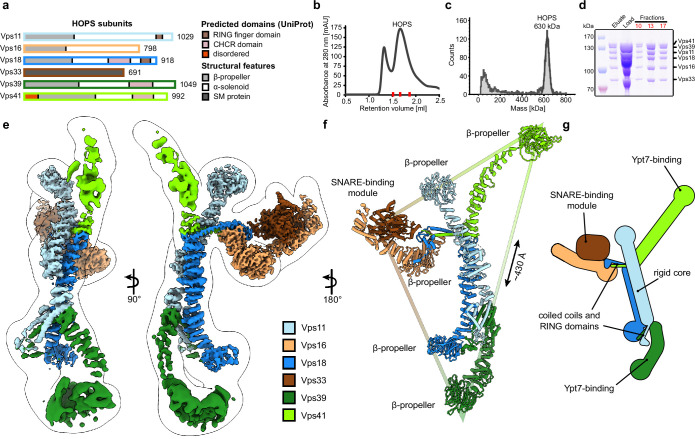
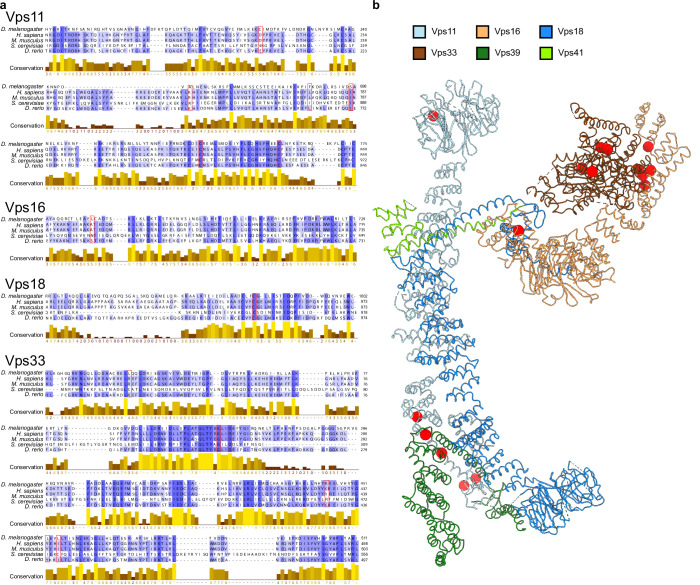
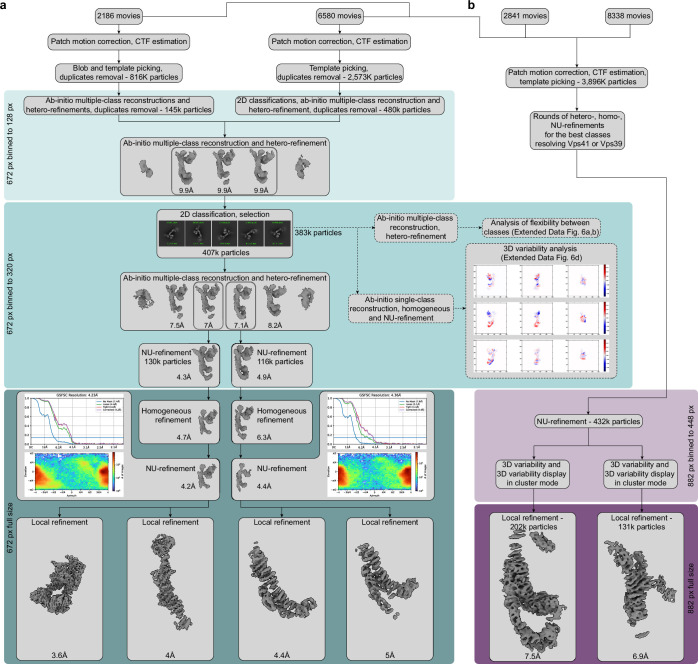
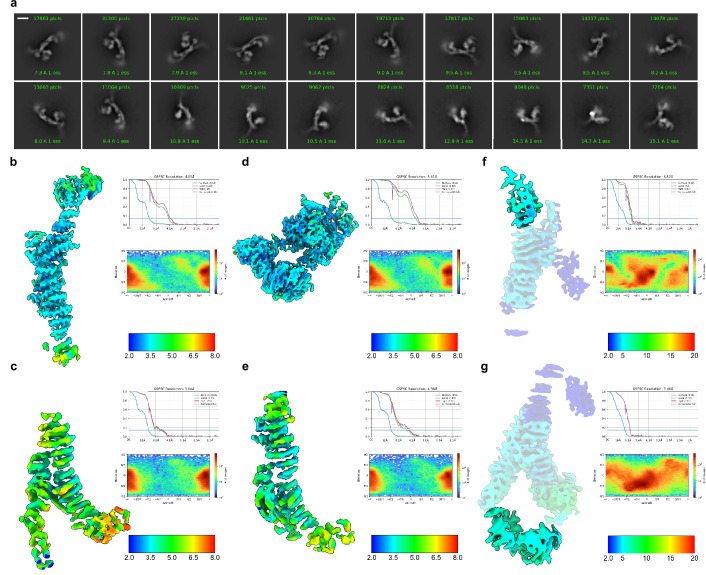
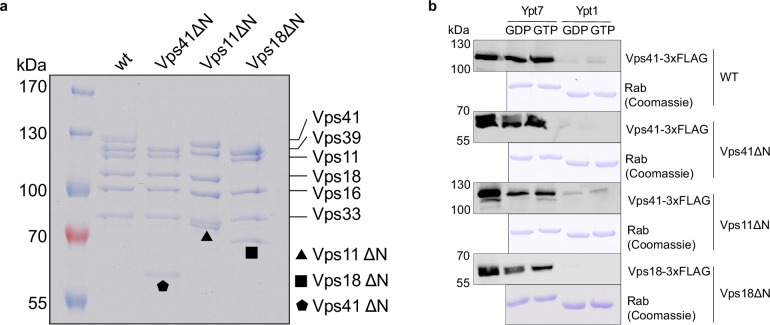
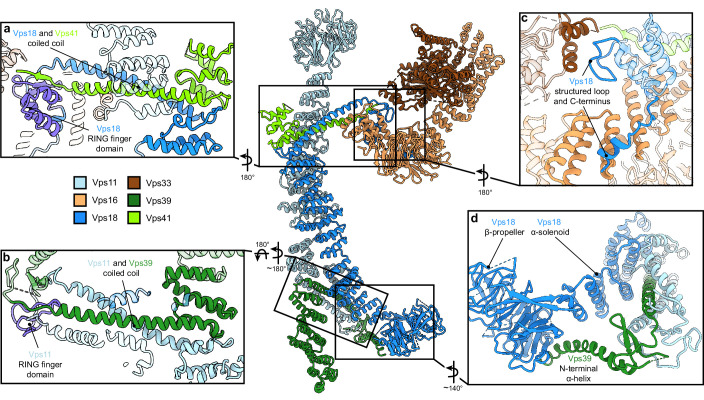
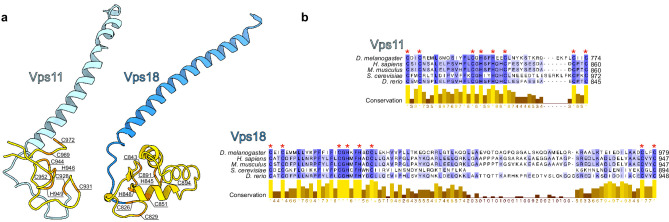
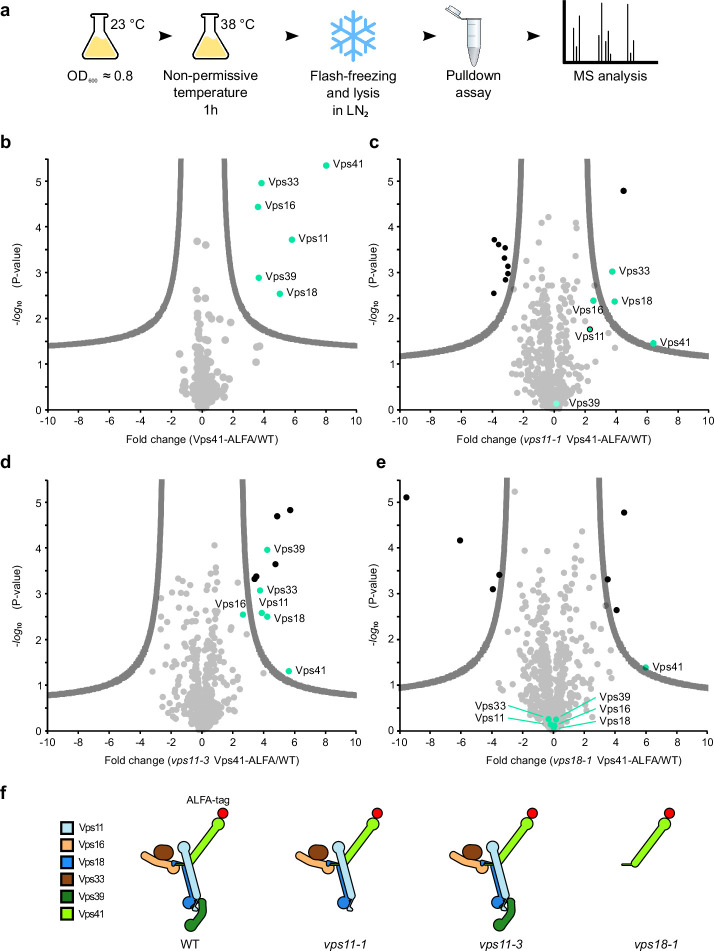
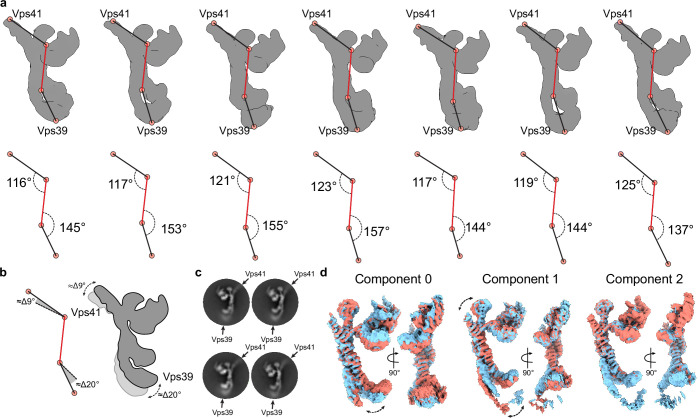
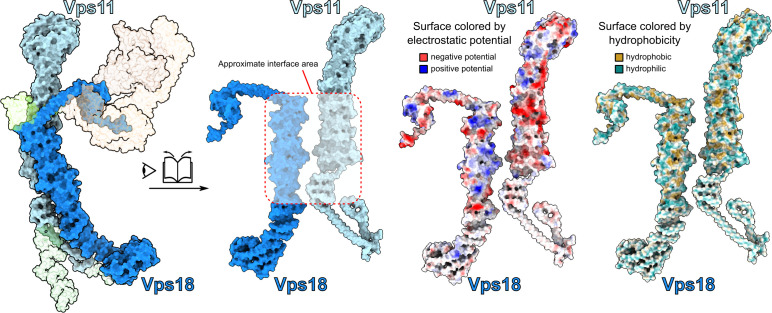
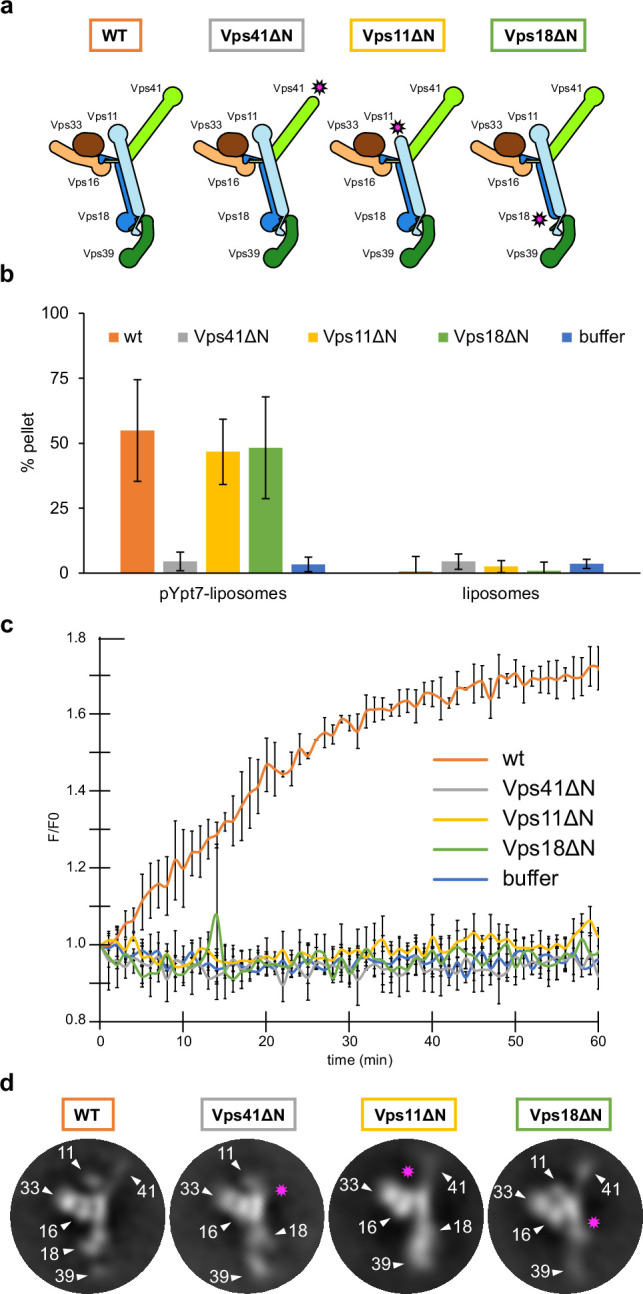
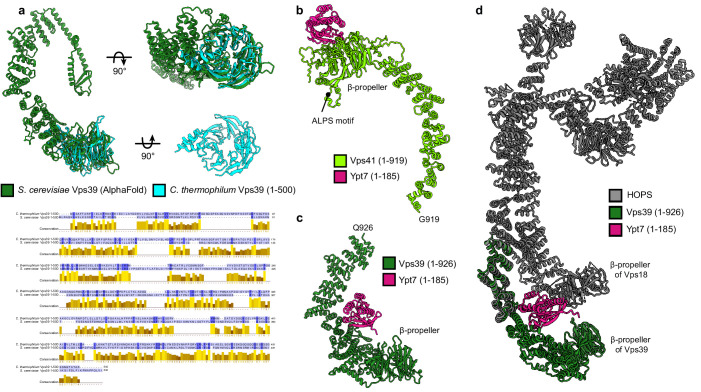
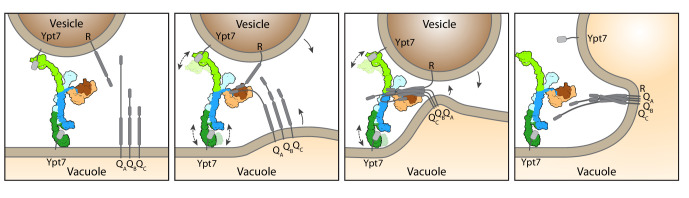
Lysosomes are essential for cellular recycling, nutrient signaling, autophagy, and pathogenic bacteria and viruses invasion. Lysosomal fusion is fundamental to cell survival and requires HOPS, a conserved heterohexameric tethering complex. On the membranes to be fused, HOPS binds small membrane-associated GTPases and assembles SNAREs for fusion, but how the complex fulfills its function remained speculative. Here, we used cryo-electron microscopy to reveal the structure of HOPS. Unlike previously reported, significant flexibility of HOPS is confined to its extremities, where GTPase binding occurs. The SNARE-binding module is firmly attached to the core, therefore, ideally positioned between the membranes to catalyze fusion. Our data suggest a model for how HOPS fulfills its dual functionality of tethering and fusion and indicate why it is an essential part of the membrane fusion machinery.
溶酶体对于细胞的再循环、营养信号、自噬以及致病性细菌和病毒的入侵都至关重要。溶酶体融合是细胞存活的基础,需要 HOPS,这是一种保守的异六聚体连接复合物。在需要融合的膜上,HOPS 结合小的膜相关 GTPase 并组装 SNARE 以进行融合,但复合物如何发挥其功能仍存在推测。在这里,我们使用冷冻电镜来揭示 HOPS 的结构。与之前报道的不同,HOPS 的显著灵活性仅限于其发生 GTPase 结合的末端。SNARE 结合模块牢固地附着在核心上,因此理想地位于膜之间以催化融合。我们的数据为 HOPS 如何发挥其连接和融合的双重功能提供了一个模型,并表明为什么它是膜融合机制的重要组成部分。