Department of Biochemistry and Biophysics, School of Medicine, University of North Carolina, Chapel Hill, United States.
Division of Chemical Biology and Medicinal Chemistry, Eshelman School of Pharmacy, University of North Carolina, Chapel Hill, United States.
Elife. 2022 Oct 6;11:e79915. doi: 10.7554/eLife.79915.
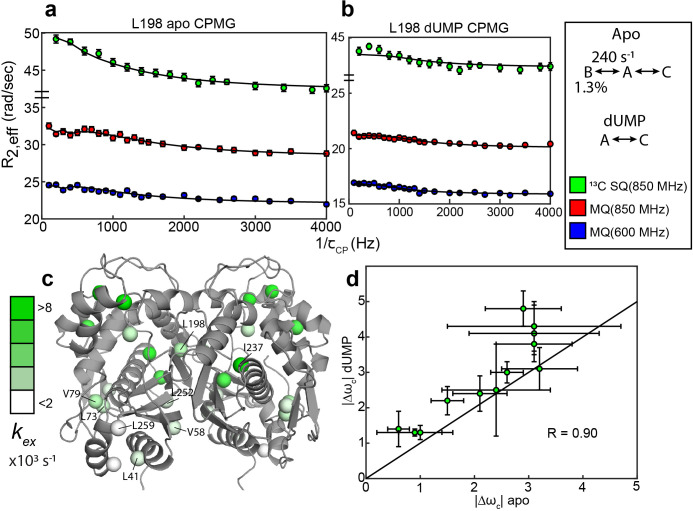
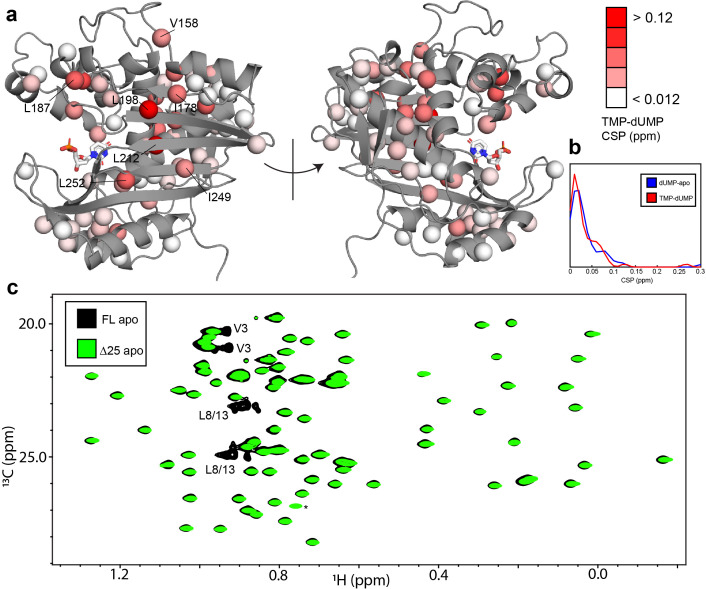
Human thymidylate synthase (hTS) is essential for DNA replication and therefore a therapeutic target for cancer. Effective targeting requires knowledge of the mechanism(s) of regulation of this 72 kDa homodimeric enzyme. Here, we investigate the mechanism of binding cooperativity of the nucleotide substrate. We have employed exquisitely sensitive methyl-based CPMG and CEST NMR experiments enabling us to identify residues undergoing bifurcated linear 3-state exchange, including concerted switching between active and inactive conformations in the apo enzyme. The inactive state is populated to only ~1.3%, indicating that conformational selection contributes negligibly to the cooperativity. Instead, methyl rotation axis order parameters, determined by H transverse relaxation rates, suggest that rigidification of the enzyme upon substrate binding is responsible for the entropically-driven cooperativity. Lack of the rigidification in product binding and substrate binding to an N-terminally truncated enzyme, both non-cooperative, support this idea. In addition, the lack of this rigidification in the N-terminal truncation indicates that interactions between the flexible N-terminus and the rest of the protein, which are perturbed by substrate binding, play a significant role in the cooperativity-a novel mechanism of dynamic allostery. Together, these findings yield a rare depth of insight into the substrate binding cooperativity of an essential enzyme.
人胸苷酸合成酶(hTS)是 DNA 复制所必需的,因此是癌症治疗的靶点。有效的靶向治疗需要了解这种 72kDa 同源二聚体酶的调节机制。在这里,我们研究了核苷酸底物结合协同性的机制。我们采用了非常灵敏的基于甲基的 CPMG 和 CEST NMR 实验,使我们能够识别经历分叉线性 3 态交换的残基,包括在apo 酶中活性和非活性构象之间的协同切换。非活性状态仅占约 1.3%,表明构象选择对协同性的贡献可以忽略不计。相反,通过 H 横向弛豫率确定的甲基旋转轴顺序参数表明,酶在底物结合时的刚性化是熵驱动协同性的原因。产物结合缺乏刚性化和底物结合到 N 端截断的酶(均无协同性)支持了这一观点。此外,N 端截断缺乏这种刚性化表明,由底物结合引起的柔性 N 端与蛋白质其余部分之间的相互作用在协同性中起着重要作用,这是一种动态变构的新机制。总之,这些发现为必需酶的底物结合协同性提供了罕见的深入见解。