School of Life Sciences, University of Dundee, Dundee, United Kingdom.
Centre for Plant Sciences, School of Biology, Faculty of Biological Sciences, University of Leeds, Leeds, United Kingdom.
Elife. 2022 Nov 21;11:e78808. doi: 10.7554/eLife.78808.
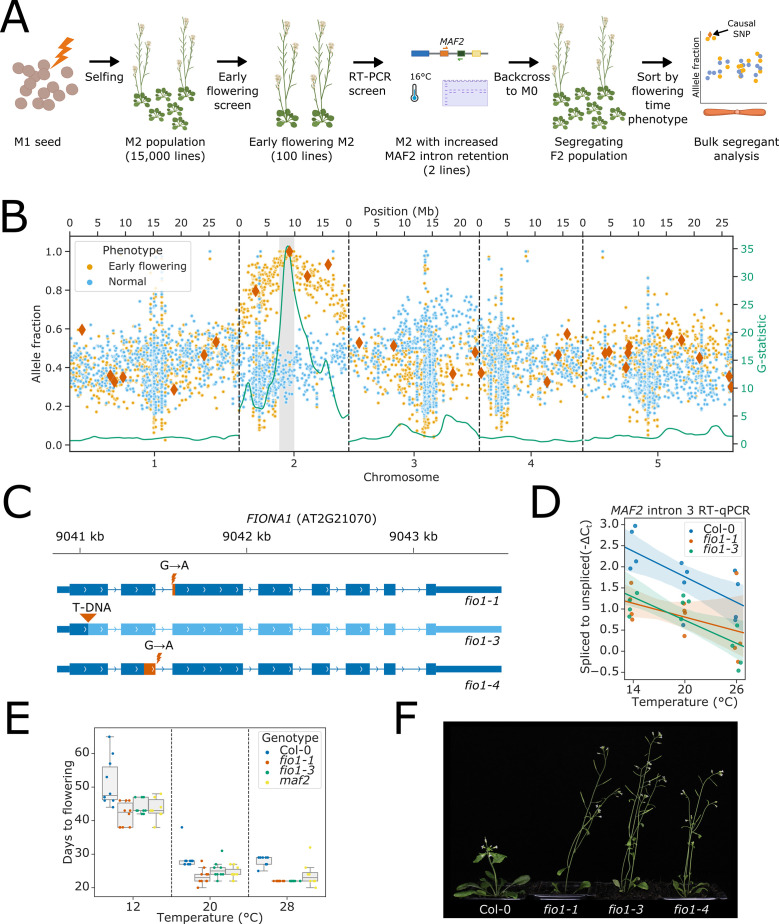
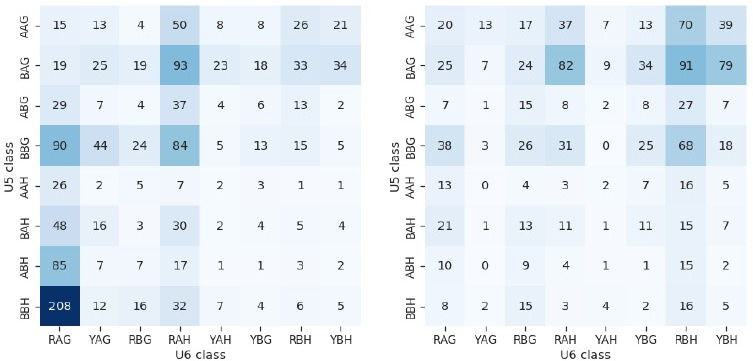
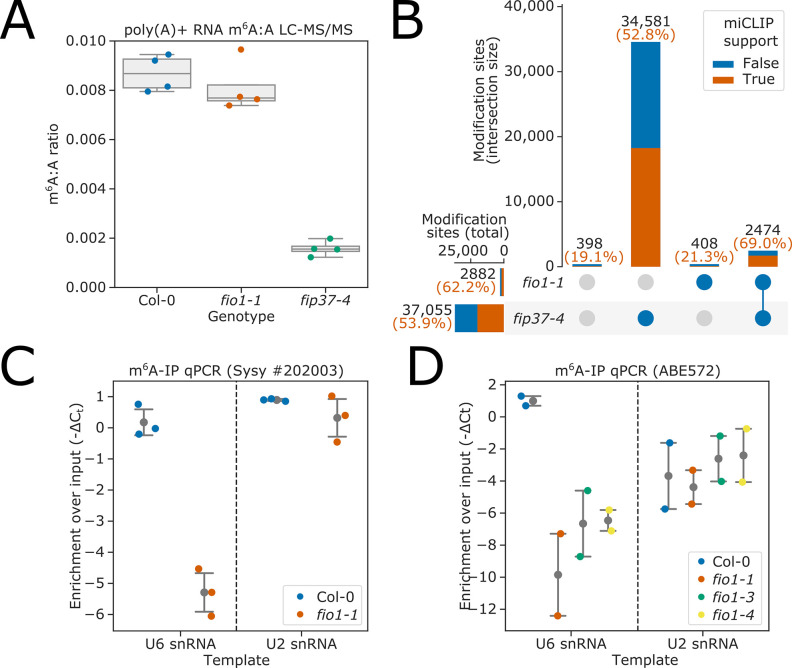
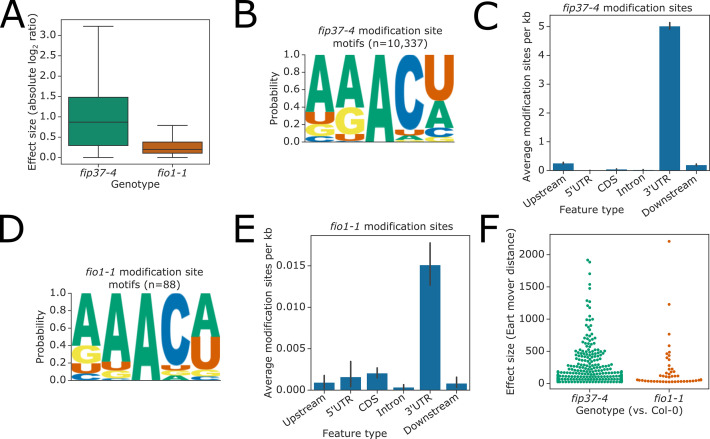
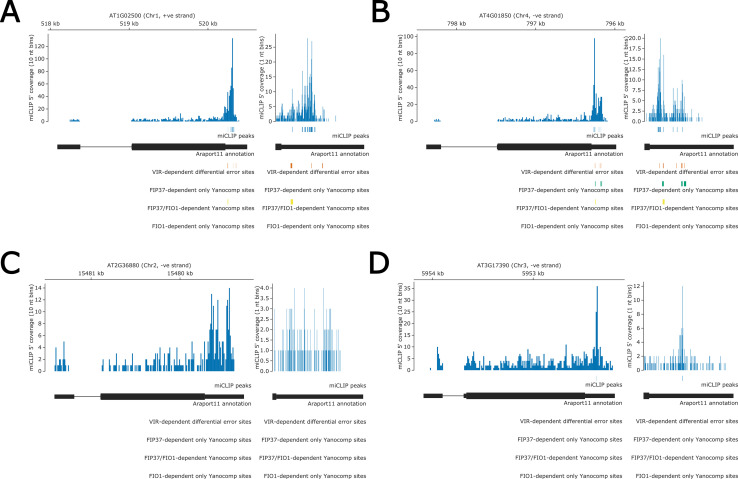
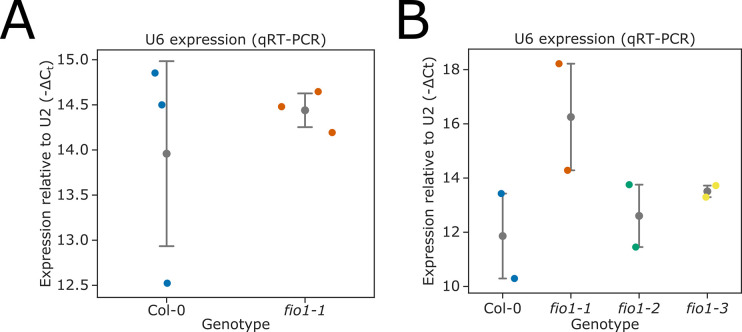
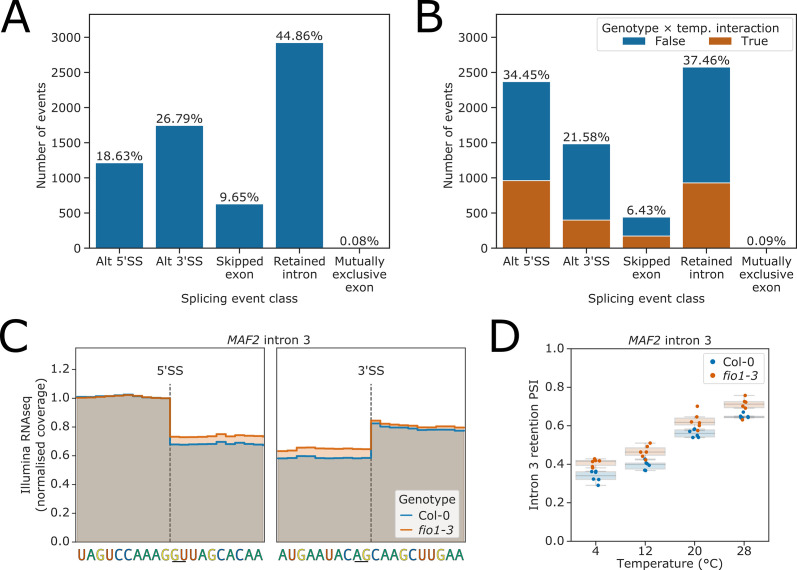
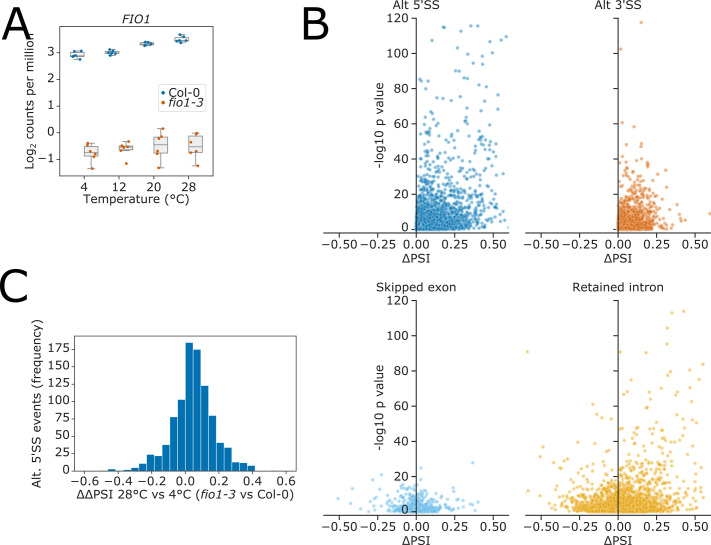
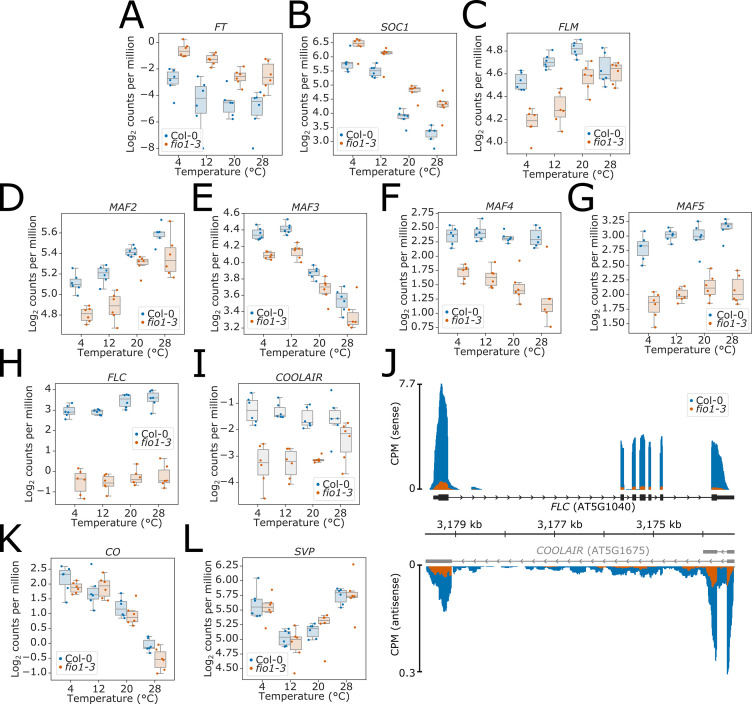
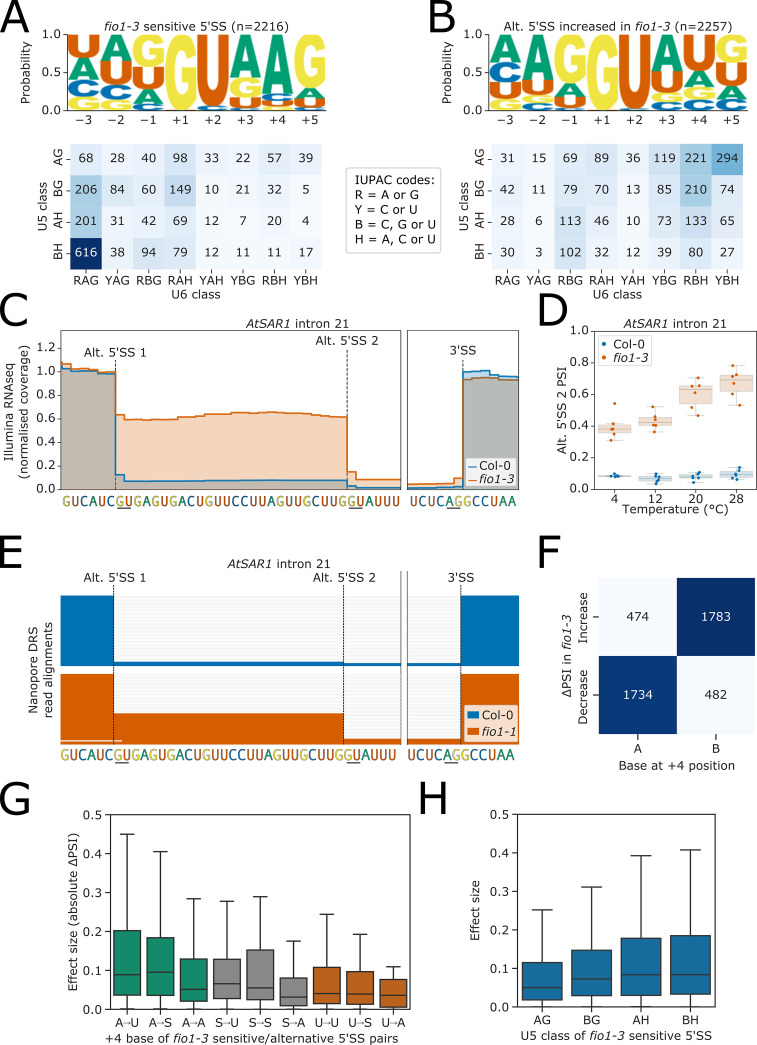
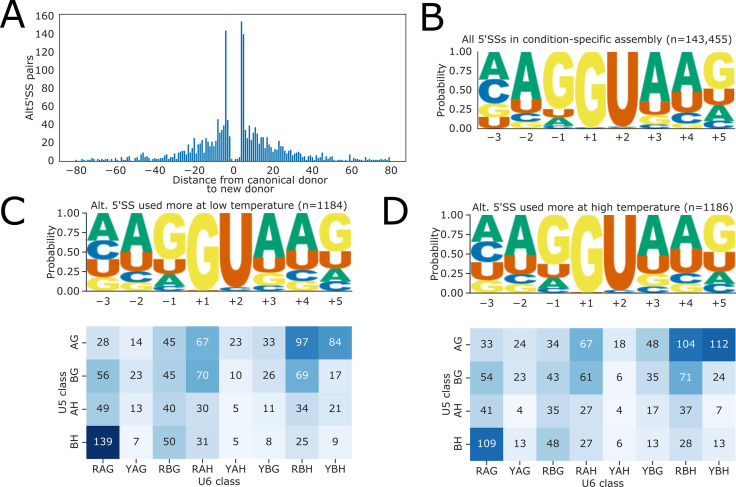
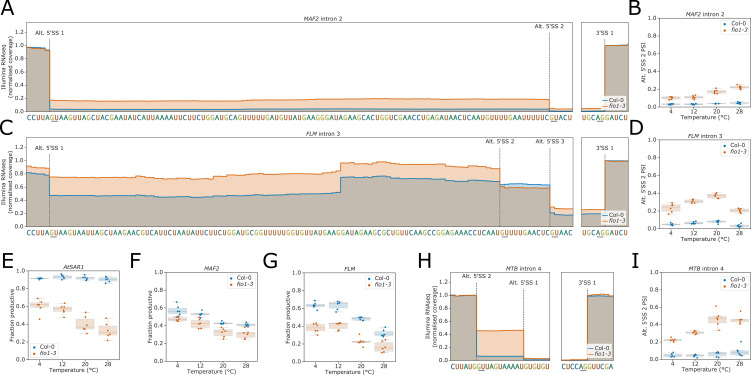
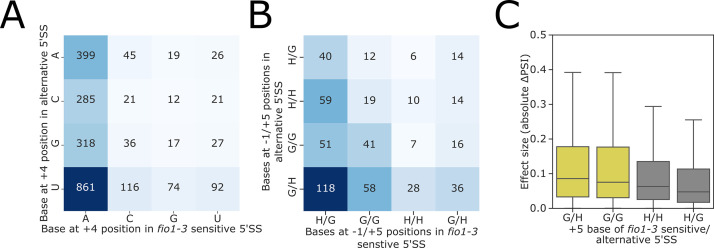
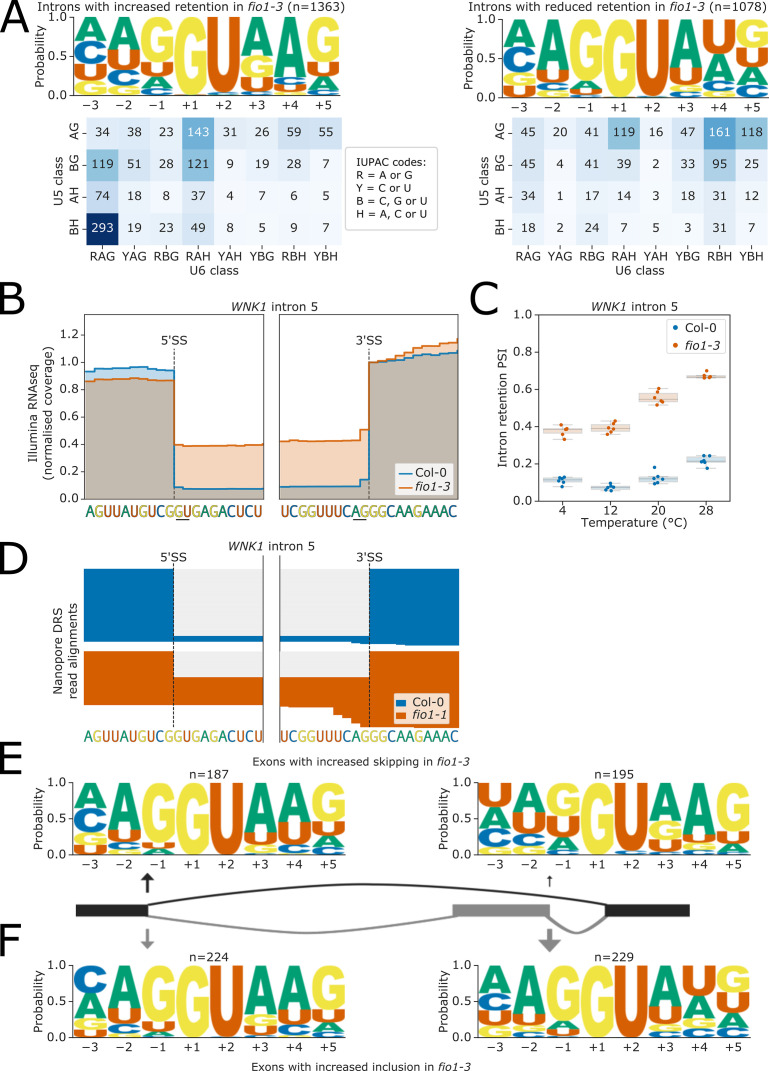
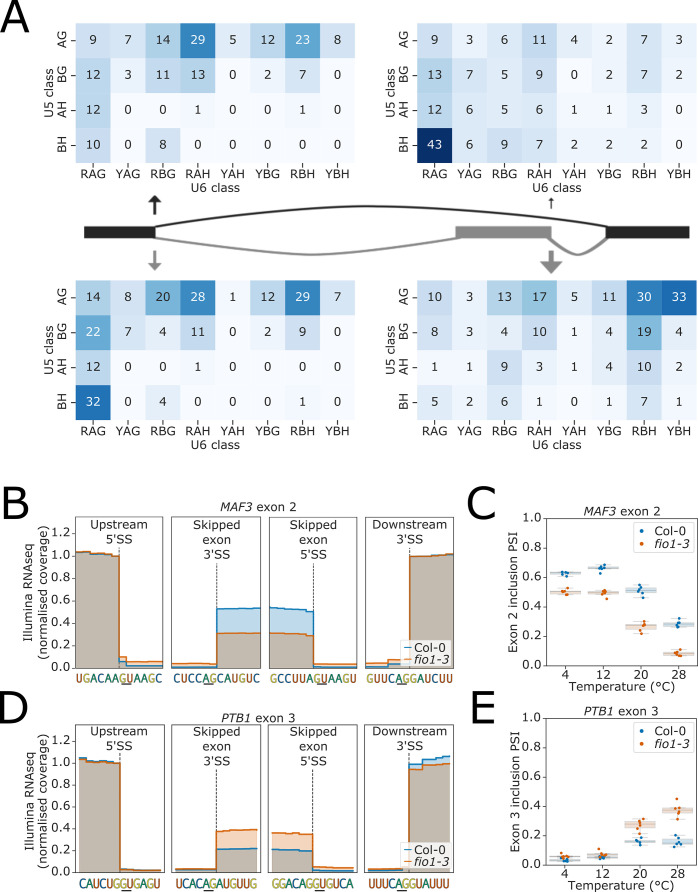
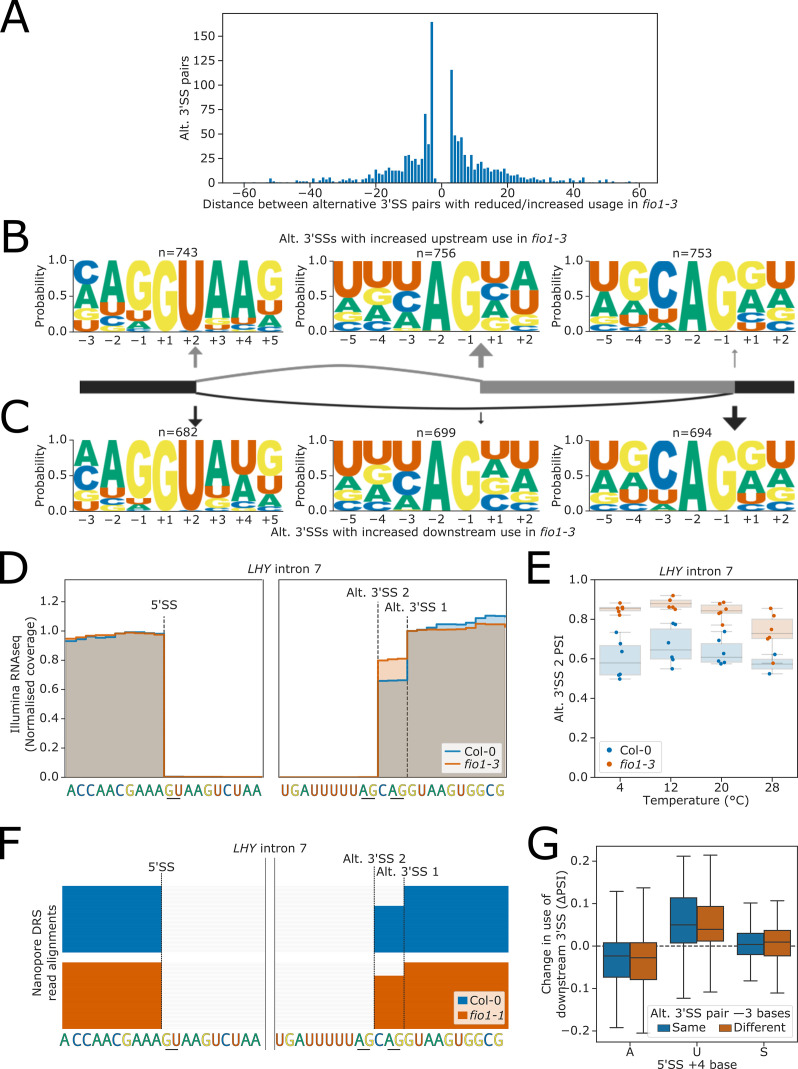
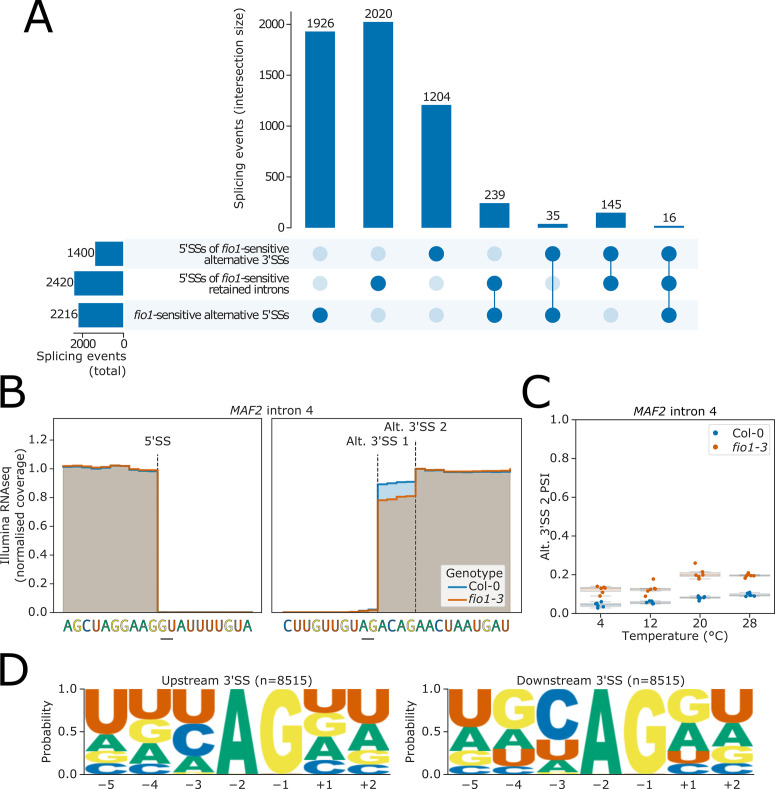
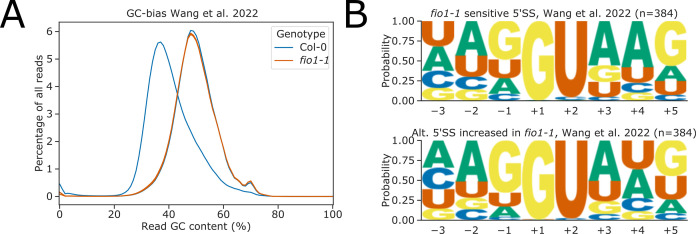
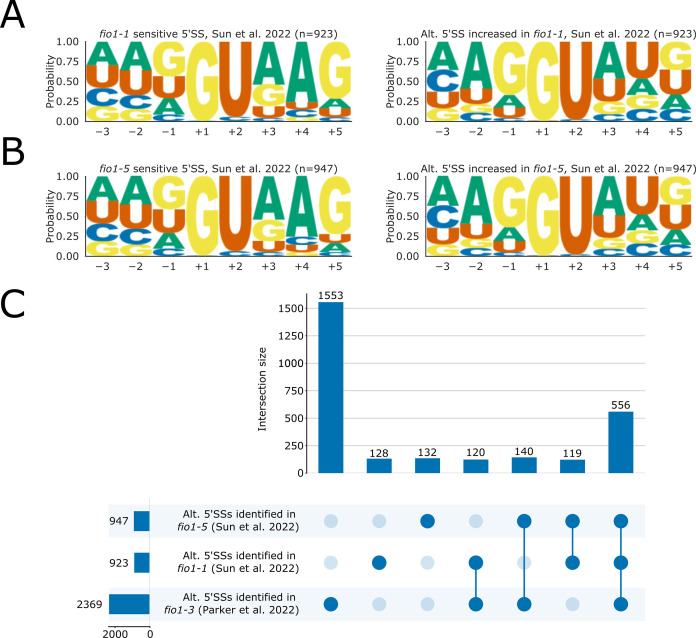
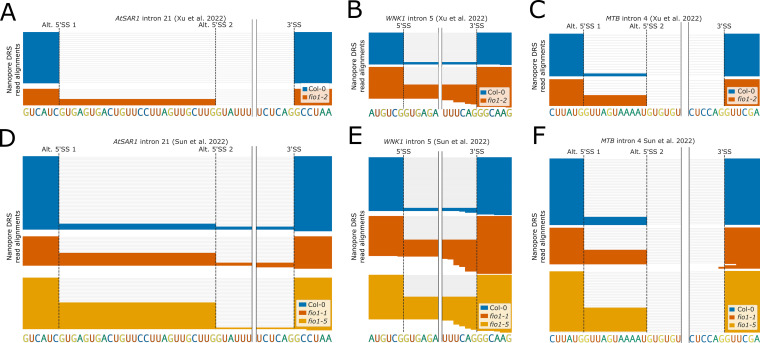
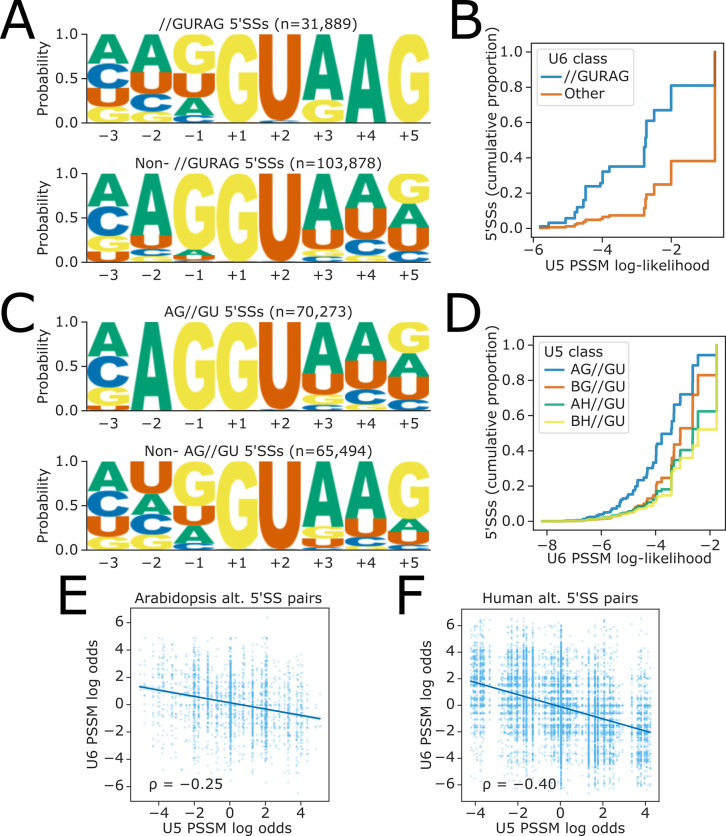
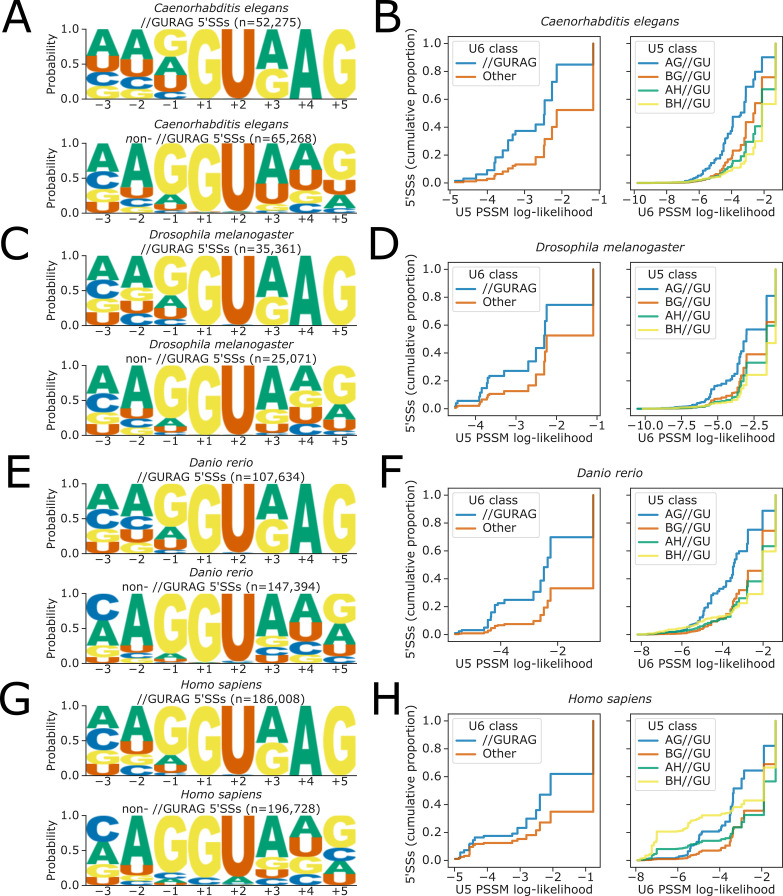
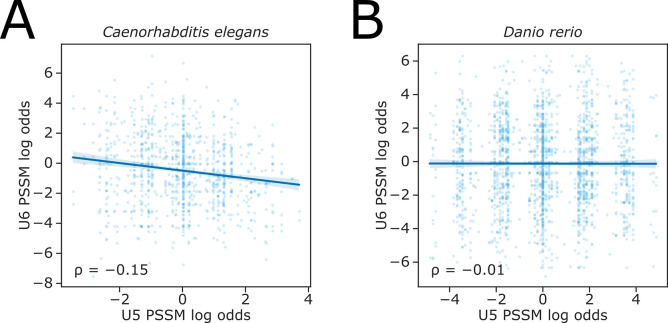
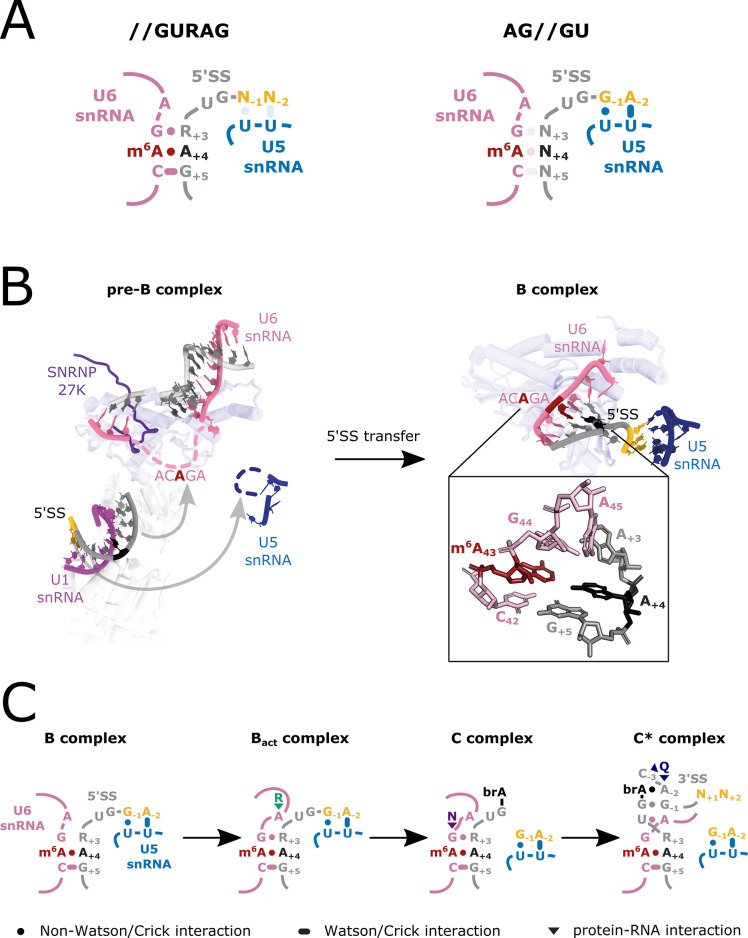
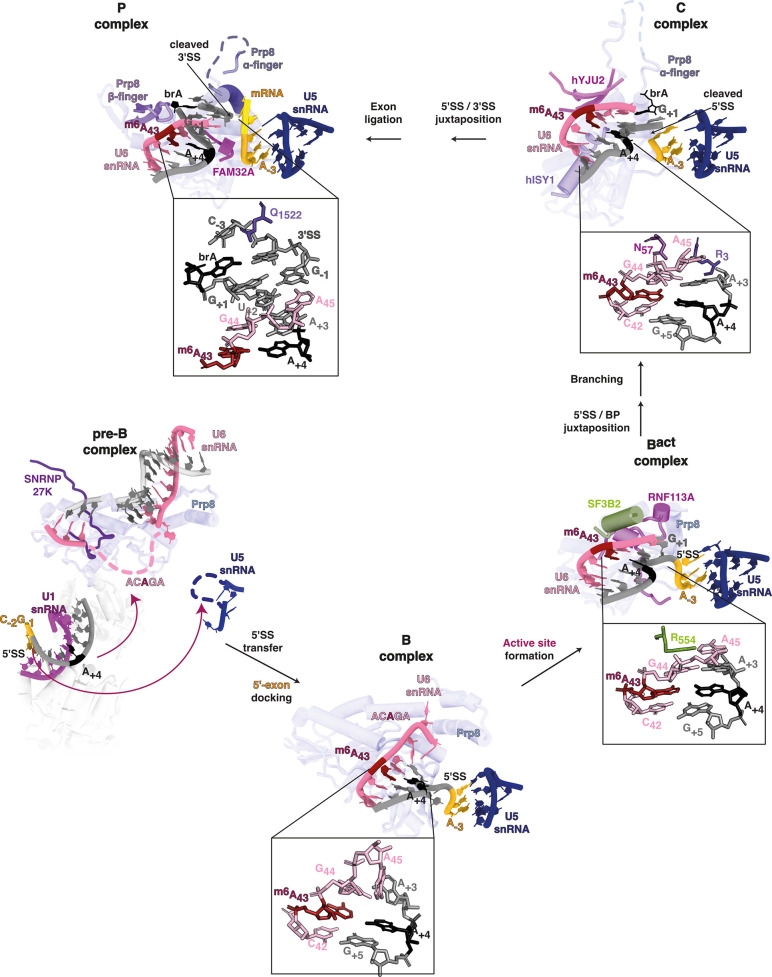
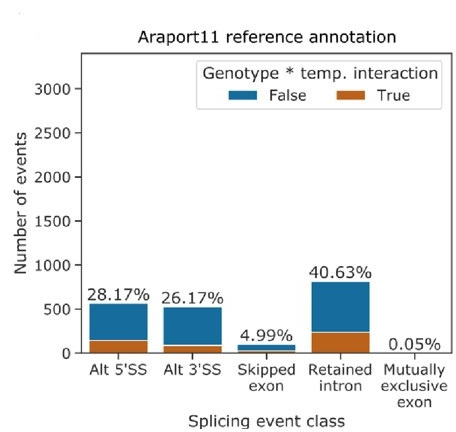
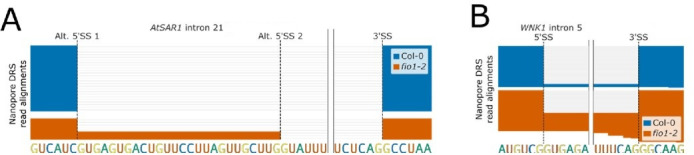
Alternative splicing of messenger RNAs is associated with the evolution of developmentally complex eukaryotes. Splicing is mediated by the spliceosome, and docking of the pre-mRNA 5' splice site into the spliceosome active site depends upon pairing with the conserved ACAGA sequence of U6 snRNA. In some species, including humans, the central adenosine of the ACGA box is modified by methylation, but the role of this mA modification is poorly understood. Here, we show that mA modified U6 snRNA determines the accuracy and efficiency of splicing. We reveal that the conserved methyltransferase, FIONA1, is required for U6 snRNA mA modification. mutants show disrupted patterns of splicing that can be explained by the sequence composition of 5' splice sites and cooperative roles for U5 and U6 snRNA in splice site selection. U6 snRNA mA influences 3' splice site usage. We generalise these findings to reveal two major classes of 5' splice site in diverse eukaryotes, which display anti-correlated interaction potential with U5 snRNA loop 1 and the U6 snRNA ACGA box. We conclude that U6 snRNA mA modification contributes to the selection of degenerate 5' splice sites crucial to alternative splicing.
信使 RNA 的可变剪接与发育复杂的真核生物的进化有关。剪接由剪接体介导,并且前体 mRNA 5'剪接位点与 U6 snRNA 的保守 ACAGA 序列配对取决于与剪接体活性位点的对接。在一些物种中,包括人类,ACGA 框的中心腺苷被甲基化修饰,但这种 mA 修饰的作用知之甚少。在这里,我们表明 mA 修饰的 U6 snRNA 决定了剪接的准确性和效率。我们揭示了保守的甲基转移酶 FIONA1 是 U6 snRNA mA 修饰所必需的。突变体显示出剪接模式的破坏,这可以通过 5'剪接位点的序列组成以及 U5 和 U6 snRNA 在剪接位点选择中的合作作用来解释。U6 snRNA mA 影响 3'剪接位点的使用。我们将这些发现推广到揭示不同真核生物中两种主要类型的 5'剪接位点,它们与 U5 snRNA 环 1 和 U6 snRNA ACGA 框显示出反相关的相互作用潜力。我们得出结论,U6 snRNA mA 修饰有助于选择对可变剪接至关重要的退化 5'剪接位点。