Department of Microbiology, Moyne Institute of Preventive Medicine, Trinity College Dublin, Dublin, Ireland.
Present address: Technical University of the Atlantic, Galway, Ireland.
Microbiology (Reading). 2023 Jan;169(1). doi: 10.1099/mic.0.001283.
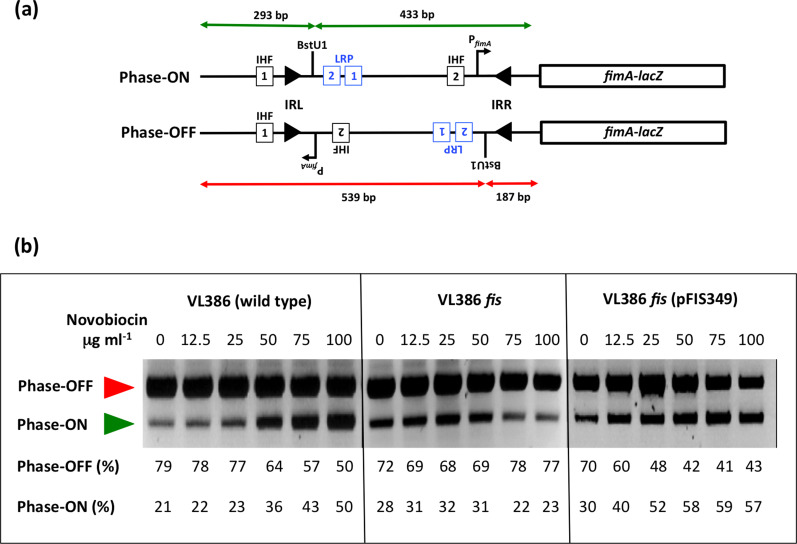
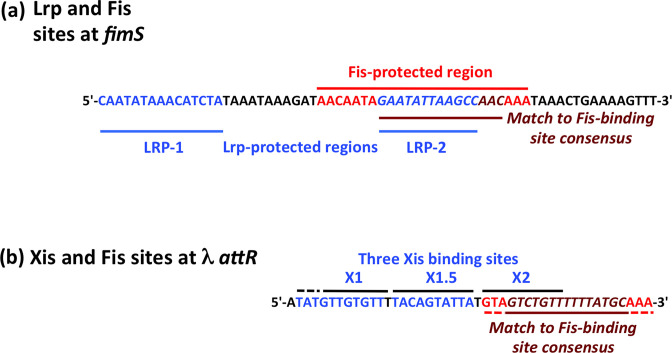
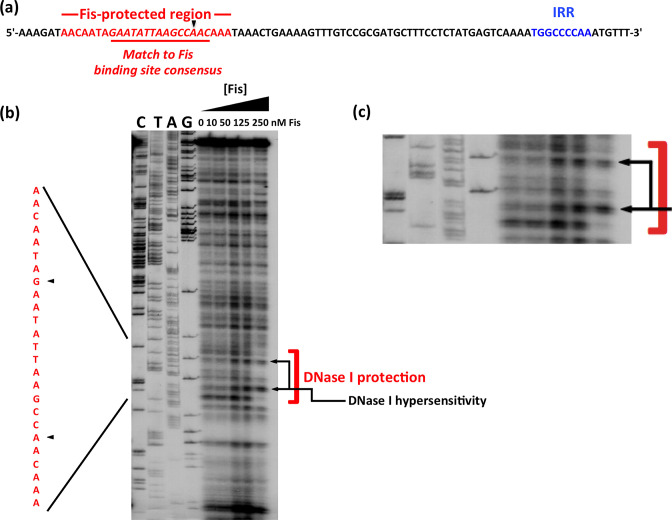
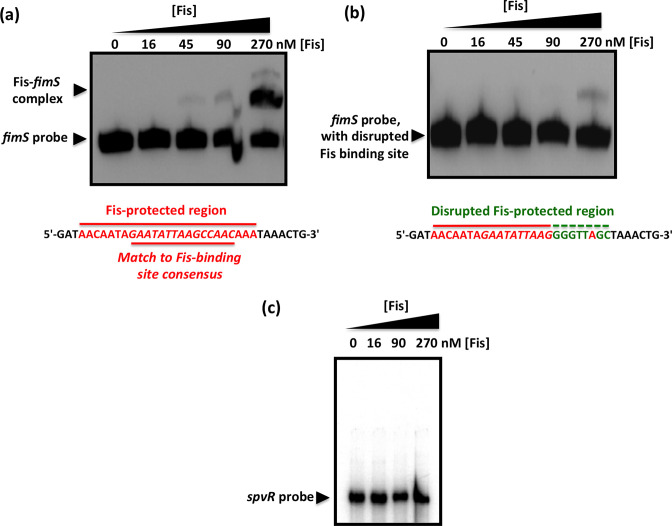
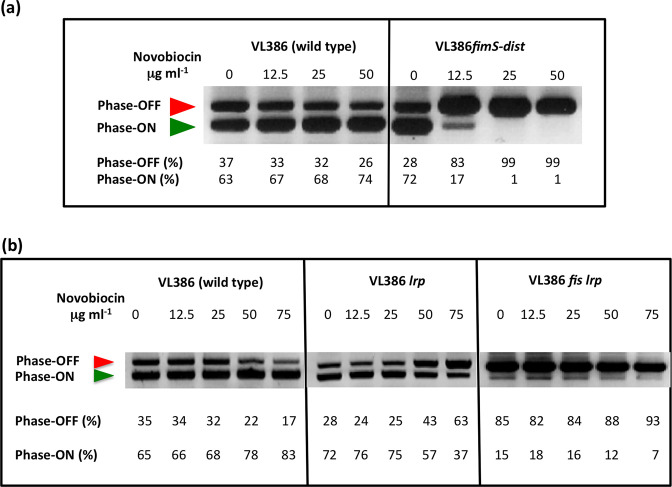
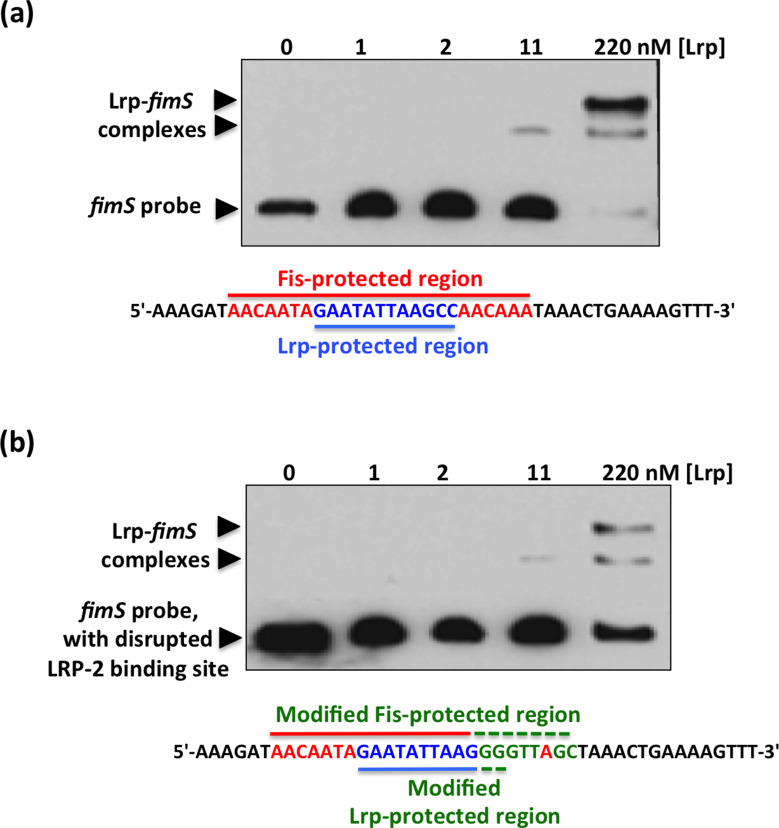
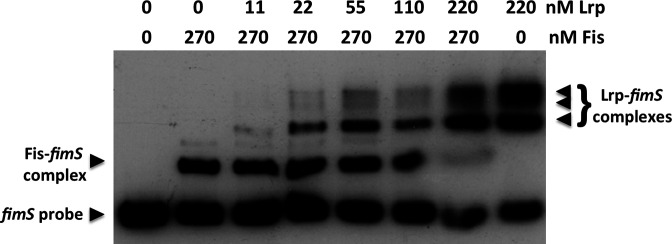
The structural genes expressing type 1 fimbriae in alternate between expressed (phase ON) and non-expressed (phase OFF) states due to inversion of the 314 bp genetic switch. The FimB tyrosine integrase inverts by site-specific recombination, alternately connecting and disconnecting the operon encoding the fimbrial subunit protein and its associated secretion and adhesin factors, to and from its transcriptional promoter within . Site-specific recombination by the FimB recombinase becomes biased towards phase ON as DNA supercoiling is relaxed, a condition that occurs when bacteria approach the stationary phase of the growth cycle. This effect can be mimicked in exponential phase cultures by inhibiting the negative DNA supercoiling activity of DNA gyrase. We report that this bias towards phase ON depends on the presence of the Fis nucleoid-associated protein. We mapped the Fis binding to a site within the invertible switch by DNase I footprinting. Disruption of this binding site by base substitution mutagenesis abolishes both Fis binding and the ability of the mutated switch to sustain its phase ON bias when DNA is relaxed, even in bacteria that produce the Fis protein. In addition, the Fis binding site overlaps one of the sites used by the Lrp protein, a known directionality determinant of inversion that also contributes to phase ON bias. The Fis–Lrp relationship at is reminiscent of that between Fis and Xis when promoting DNA relaxation-dependent excision of bacteriophage λ from the chromosome. However, unlike the co-binding mechanism used by Fis and Xis at λ , the Fis–Lrp relationship at involves competitive binding. We discuss these findings in the context of the link between inversion biasing and the physiological state of the bacterium.
表达 1 型菌毛的结构基因在表达(ON 相)和非表达(OFF 相)状态之间交替,这是由于 314bp 遗传开关的倒置。FimB 酪氨酸整合酶通过位点特异性重组进行倒置,交替连接和断开编码菌毛亚基蛋白及其相关分泌和粘附因子的操纵子,与转录启动子 within 连接或断开。FimB 重组酶的位点特异性重组在 DNA 超螺旋松弛时偏向于 ON 相,这种情况发生在细菌接近生长周期的静止期时。在指数生长期培养物中,通过抑制 DNA 拓扑异构酶的负 DNA 超螺旋活性,可以模拟这种偏向 ON 相的效应。我们报告称,这种偏向 ON 相取决于 Fis 核蛋白的存在。我们通过 DNase I 足迹法将 Fis 结合定位到可反转的开关内的一个位点。通过碱基取代诱变破坏该结合位点,不仅会破坏 Fis 结合,还会破坏突变开关在 DNA 松弛时维持其 ON 相偏向的能力,即使在产生 Fis 蛋白的细菌中也是如此。此外,Fis 结合位点重叠了 Lrp 蛋白使用的一个位点之一,Lrp 蛋白是 倒置的已知方向性决定因素,也有助于 ON 相偏向。在 处的 Fis-Lrp 关系类似于促进噬菌体 λ从 染色体上 DNA 松弛依赖性切除时 Fis 和 Xis 之间的关系。然而,与 Fis 和 Xis 在 λ 上使用的共结合机制不同,Fis-Lrp 关系在 处涉及竞争结合。我们在细菌生理状态与 倒置偏倚之间的联系的背景下讨论这些发现。