Department of Experimental Radiation Oncology, The University of Texas MD Anderson Cancer Center, Houston, TX, 77030, USA.
Kadmon Corporation, LLC (A Sanofi Company), New York, NY, 10016, USA.
Nat Commun. 2023 Jun 21;14(1):3673. doi: 10.1038/s41467-023-39401-9.
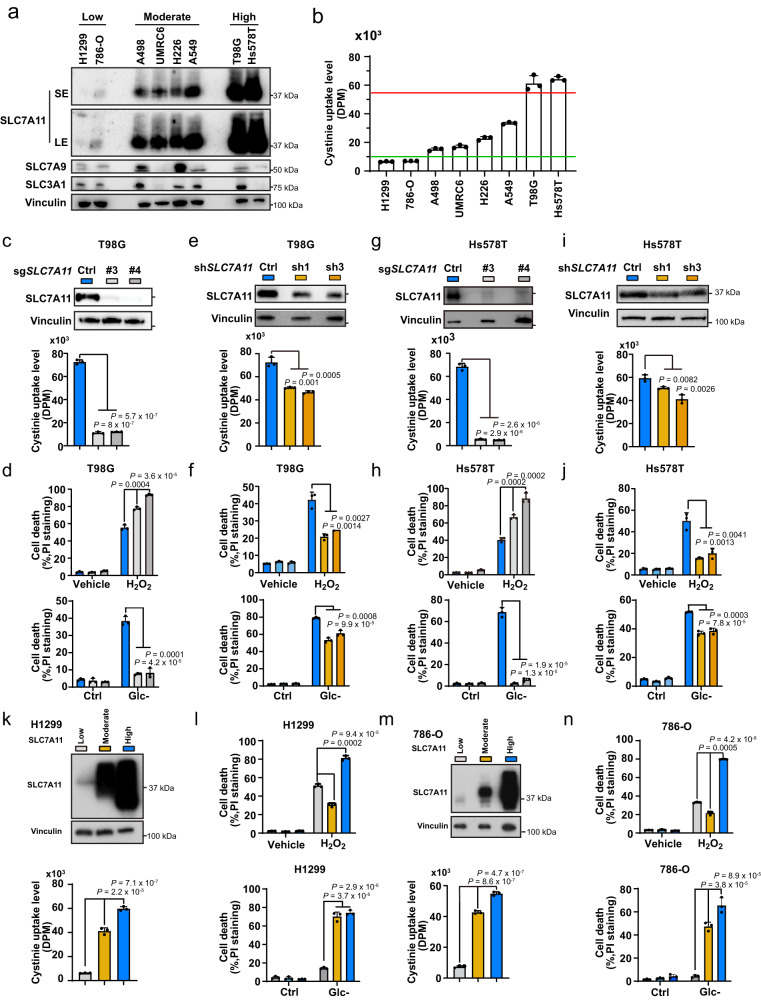
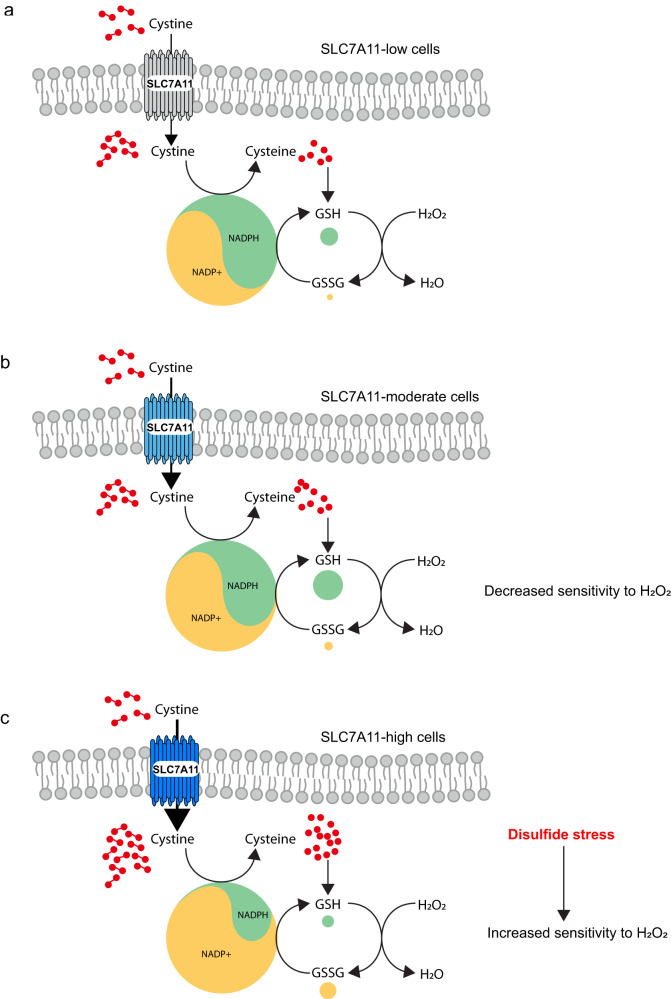
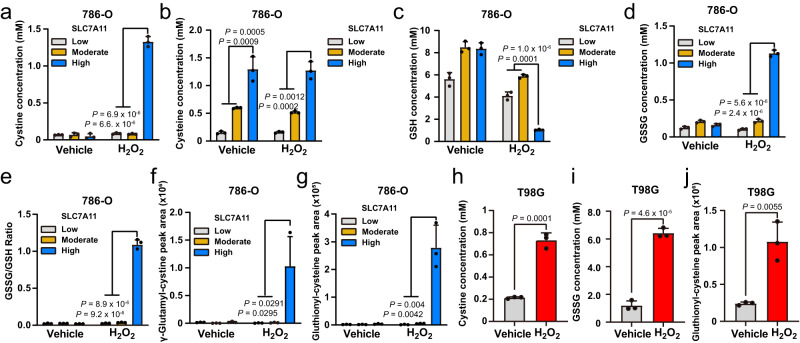
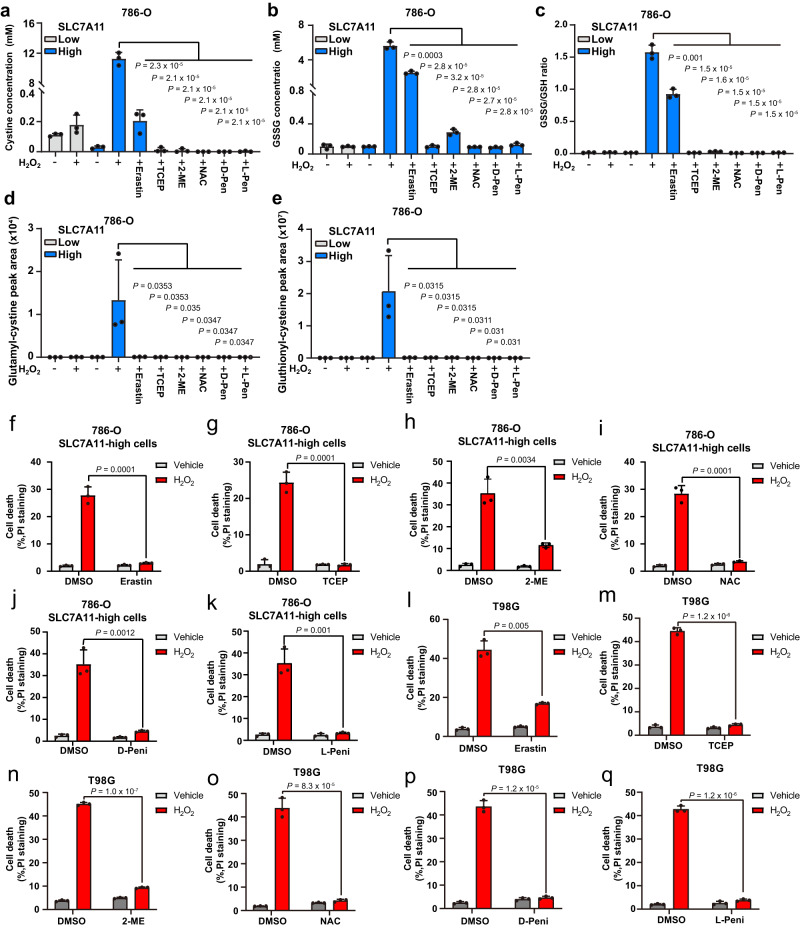
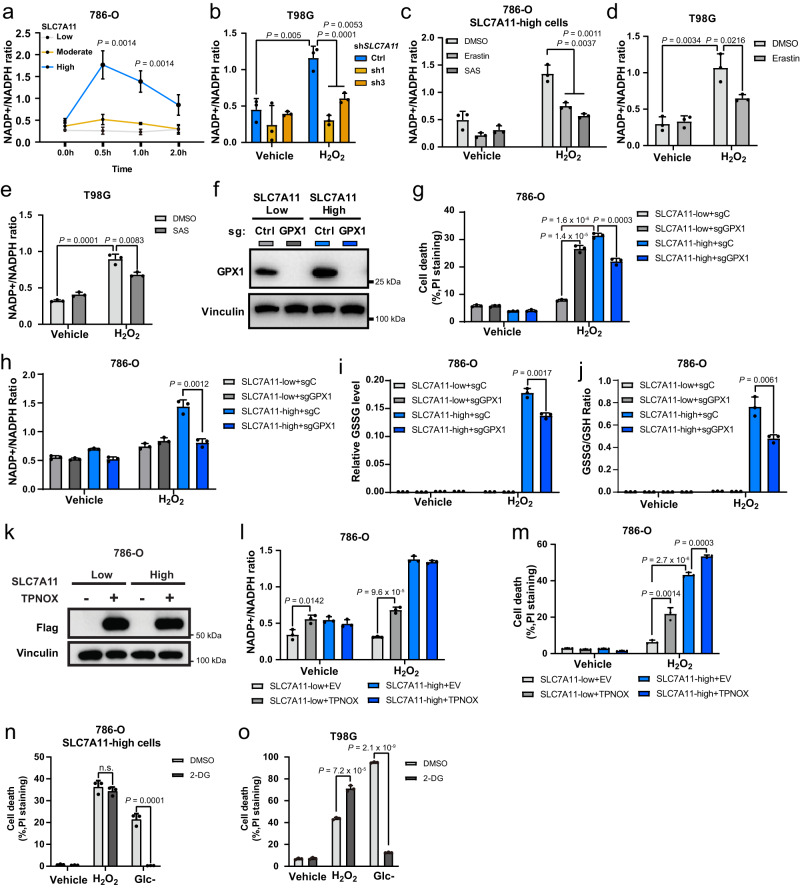
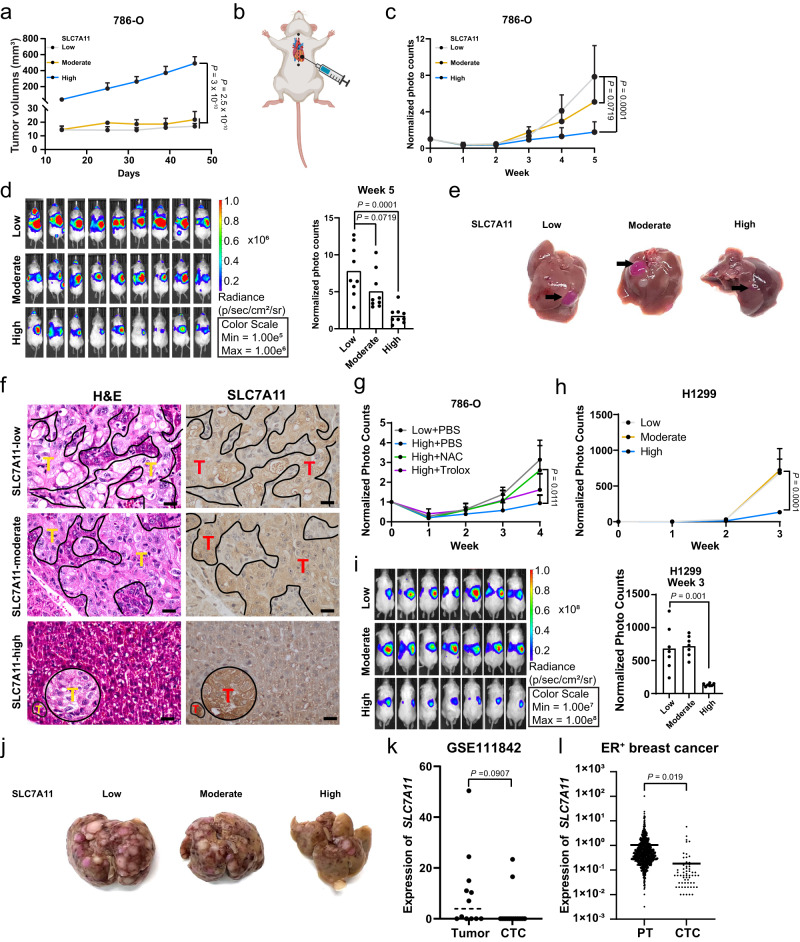
The cystine transporter solute carrier family 7 member 11 (SLC7A11; also called xCT) protects cancer cells from oxidative stress and is overexpressed in many cancers. Here we report a surprising finding that, whereas moderate overexpression of SLC7A11 is beneficial for cancer cells treated with HO, a common oxidative stress inducer, its high overexpression dramatically increases HO-induced cell death. Mechanistically, high cystine uptake in cancer cells with high overexpression of SLC7A11 in combination with HO treatment results in toxic buildup of intracellular cystine and other disulfide molecules, NADPH depletion, redox system collapse, and rapid cell death (likely disulfidptosis). We further show that high overexpression of SLC7A11 promotes tumor growth but suppresses tumor metastasis, likely because metastasizing cancer cells with high expression of SLC7A11 are particularly susceptible to oxidative stress. Our findings reveal that SLC7A11 expression level dictates cancer cells' sensitivity to oxidative stress and suggests a context-dependent role for SLC7A11 in tumor biology.
胱氨酸转运蛋白溶质载体家族 7 成员 11(SLC7A11;也称为 xCT)可保护癌细胞免受氧化应激,并且在许多癌症中过度表达。在这里,我们报告了一个令人惊讶的发现,即 SLC7A11 的适度过表达有利于用 HO(一种常见的氧化应激诱导剂)处理的癌细胞,但高过表达会显著增加 HO 诱导的细胞死亡。从机制上讲,在 SLC7A11 高过表达的癌细胞中,高胱氨酸摄取与 HO 处理相结合,导致细胞内胱氨酸和其他二硫分子的毒性积累、NADPH 耗竭、氧化还原系统崩溃和快速细胞死亡(可能是二硫细胞死亡)。我们进一步表明,SLC7A11 的高过表达促进肿瘤生长,但抑制肿瘤转移,可能是因为高表达 SLC7A11 的转移癌细胞特别容易受到氧化应激的影响。我们的发现揭示了 SLC7A11 的表达水平决定了癌细胞对氧化应激的敏感性,并提示 SLC7A11 在肿瘤生物学中具有上下文相关的作用。