Protein Processing Section, Structural Biophysics Laboratory, Center for Cancer Research, National Cancer Institute, National Institutes of Health, Frederick, Maryland, USA.
Genome Modification Core, Frederick National Laboratory for Cancer Research, Frederick, Maryland, USA.
J Biol Chem. 2023 Aug;299(8):104948. doi: 10.1016/j.jbc.2023.104948. Epub 2023 Jun 22.
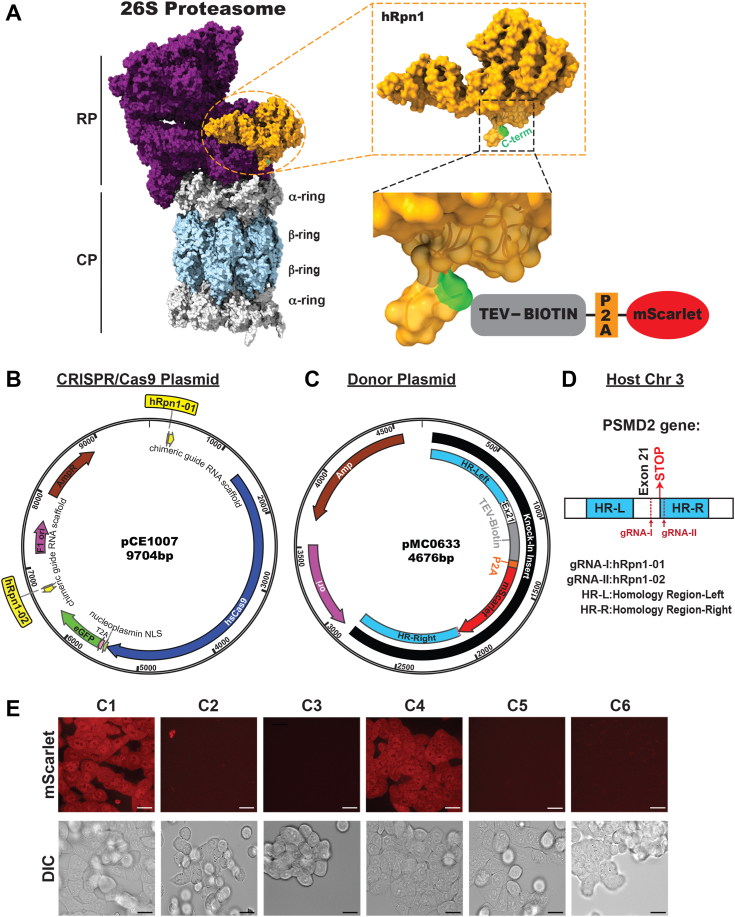
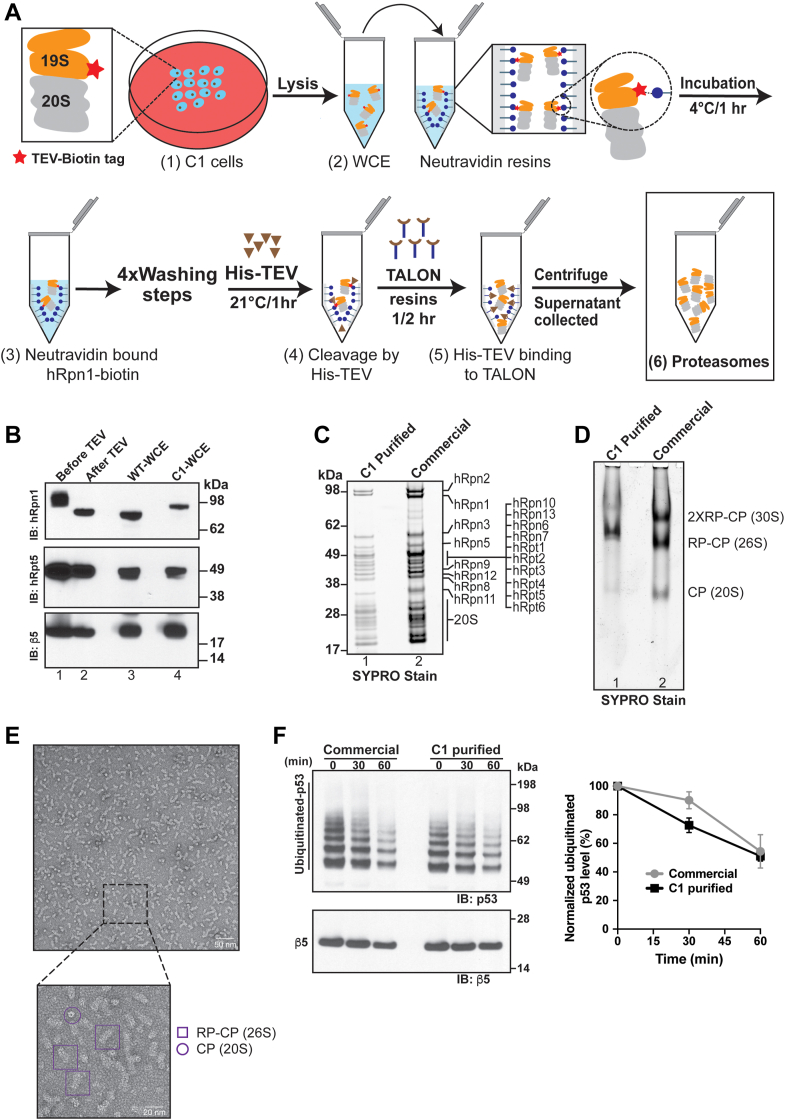
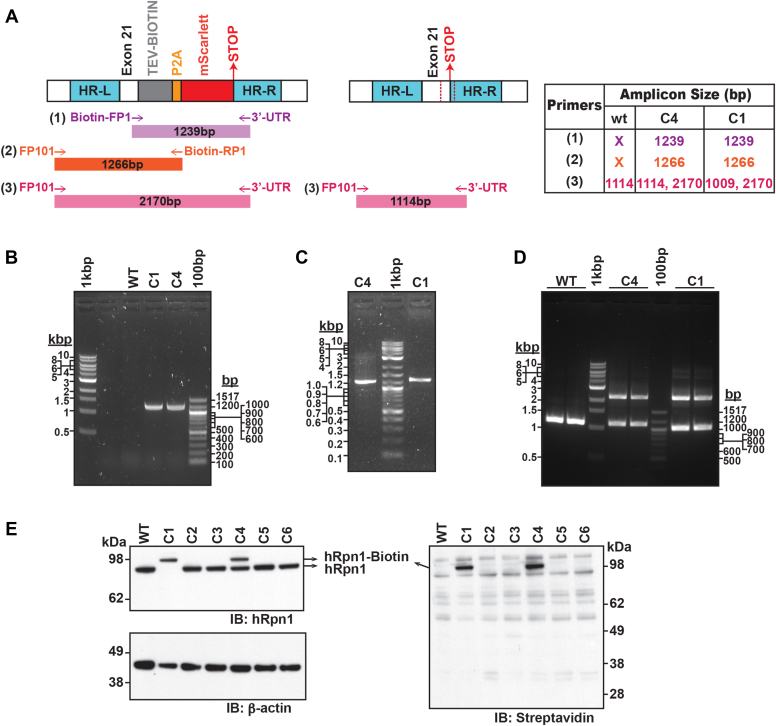
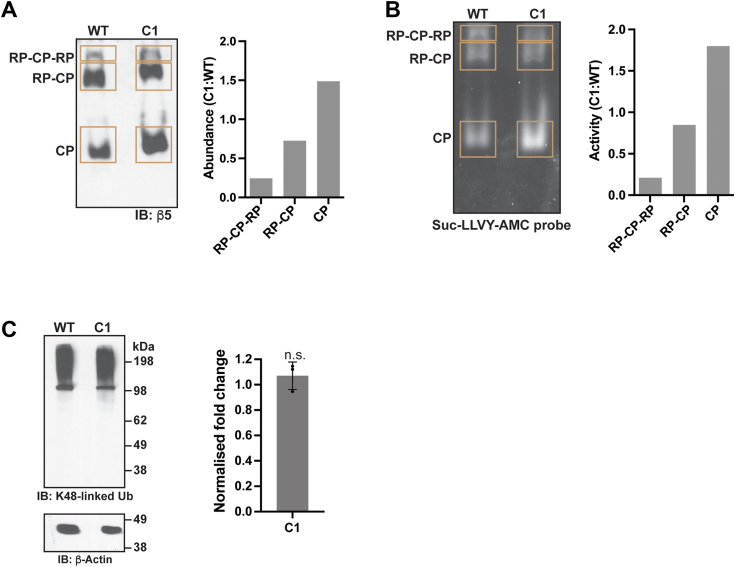
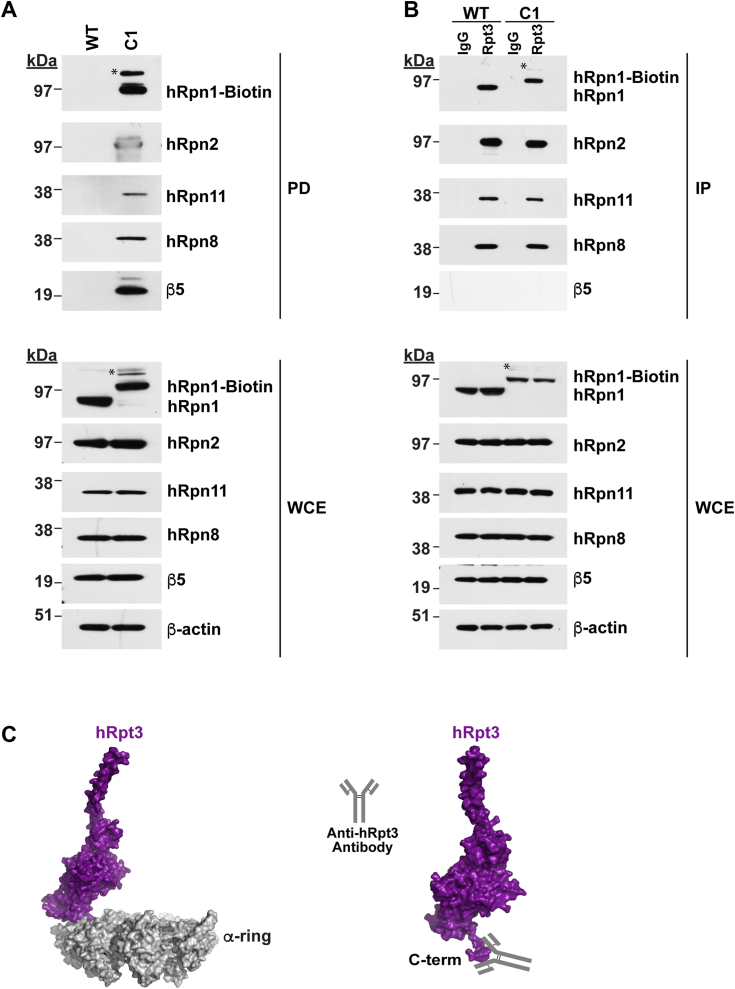
Regulated protein degradation in eukaryotes is performed by the 26S proteasome, which contains a 19-subunit regulatory particle (RP) that binds, processes, and translocates substrates to a 28-subunit hollow core particle (CP) where proteolysis occurs. In addition to its intrinsic subunits, myriad proteins interact with the proteasome transiently, including factors that assist and/or regulate its degradative activities. Efforts to identify proteasome-interacting components and/or to solve its structure have relied on over-expression of a tagged plasmid, establishing stable cell lines, or laborious purification protocols to isolate native proteasomes from cells. Here, we describe an engineered human cell line, derived from colon cancer HCT116 cells, with a biotin handle on the RP subunit hRpn1/PSMD2 (proteasome 26S subunit, non-ATPase 2) for purification of 26S proteasomes. A 75-residue sequence from Propionibacterium shermanii that is biotinylated in mammalian cells was added following a tobacco etch virus protease cut site at the C terminus of hRpn1. We tested and found that 26S proteasomes can be isolated from this modified HCT116 cell line by using a simple purification protocol. More specifically, biotinylated proteasomes were purified from the cell lysates by using neutravidin agarose resin and released from the resin following incubation with tobacco etch virus protease. The purified proteasomes had equivalent activity in degrading a model ubiquitinated substrate, namely ubiquitinated p53, compared to commercially available bovine proteasomes that were purified by fractionation. In conclusion, advantages of this approach to obtain 26S proteasomes over others is the simple purification protocol and that all cellular proteins, including the tagged hRpn1 subunit, remain at endogenous stoichiometry.
真核生物中的调节蛋白降解是由 26S 蛋白酶体完成的,该蛋白酶体包含一个由 19 个亚基组成的调节颗粒(RP),该颗粒结合、加工和转运底物到一个由 28 个亚基组成的空心核心颗粒(CP)中,在那里发生蛋白水解。除了其内在的亚基外,无数的蛋白质与蛋白酶体短暂相互作用,包括协助和/或调节其降解活性的因子。鉴定蛋白酶体相互作用成分和/或解决其结构的努力依赖于标记质粒的过表达、建立稳定的细胞系或费力的纯化方案,从细胞中分离天然蛋白酶体。在这里,我们描述了一种源自结肠癌 HCT116 细胞的工程化人细胞系,该细胞系在 RP 亚基 hRpn1/PSMD2(蛋白酶体 26S 亚基,非 ATP 酶 2)上带有生物素手柄,用于纯化 26S 蛋白酶体。烟草蚀纹病毒蛋白酶切割位点位于 hRpn1 的 C 末端后,在哺乳动物细胞中生物素化的短杆菌肽 S 来自 Propionibacterium shermanii 的 75 个残基序列。我们进行了测试,发现可以通过使用简单的纯化方案从该修饰的 HCT116 细胞系中分离 26S 蛋白酶体。更具体地说,通过使用链霉亲和素琼脂糖树脂从细胞裂解物中纯化生物素化的蛋白酶体,并在用烟草蚀纹病毒蛋白酶孵育后从树脂中释放。与通过分级分离纯化的商业上可获得的牛蛋白酶体相比,纯化的蛋白酶体在降解模型泛素化底物(即泛素化 p53)方面具有同等的活性。总之,与其他方法相比,这种获得 26S 蛋白酶体的方法的优点是简单的纯化方案以及所有细胞蛋白,包括标记的 hRpn1 亚基,都保持内源性比例。