Department of Biology and Ecology, University of Ostrava, 710 00 Ostrava, Czech Republic.
Emeritus Laboratory of Cellular Dynamics, Max Planck Institute for Multidisciplinary Sciences, 37077 Göttingen, Germany.
Int J Mol Sci. 2023 Jun 27;24(13):10740. doi: 10.3390/ijms241310740.
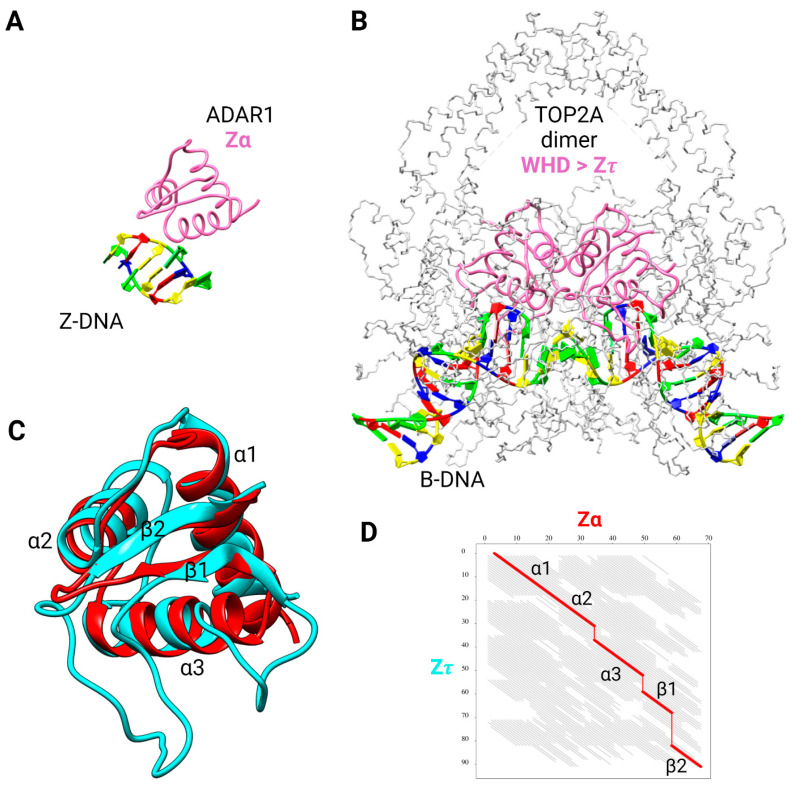
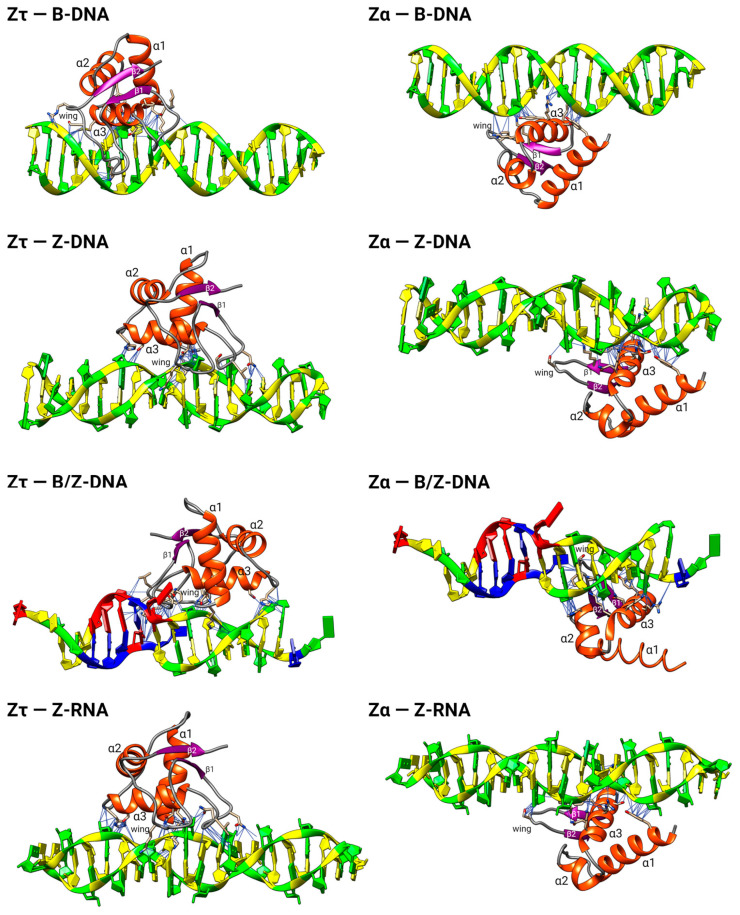
The dynamic processes operating on genomic DNA, such as gene expression and cellular division, lead inexorably to topological challenges in the form of entanglements, catenanes, knots, "bubbles", R-loops, and other outcomes of supercoiling and helical disruption. The resolution of toxic topological stress is the function attributed to DNA topoisomerases. A prominent example is the negative supercoiling (nsc) trailing processive enzymes such as DNA and RNA polymerases. The multiple equilibrium states that nscDNA can adopt by redistribution of helical twist and writhe include the left-handed double-helical conformation known as Z-DNA. Thirty years ago, one of our labs isolated a protein from cells and embryos with a 100-fold greater affinity for Z-DNA than for B-DNA, and identified it as topoisomerase II (gene Top2, orthologous to the human UniProt proteins TOP2A and TOP2B). GTP increased the affinity and selectivity for Z-DNA even further and also led to inhibition of the isomerase enzymatic activity. An allosteric mechanism was proposed, in which topoII acts as a Z-DNA-binding protein (ZBP) to stabilize given states of topological (sub)domains and associated multiprotein complexes. We have now explored this possibility by comprehensive bioinformatic analyses of the available protein sequences of topoII representing organisms covering the whole tree of life. Multiple alignment of these sequences revealed an extremely high level of evolutionary conservation, including a winged-helix protein segment, here denoted as Zτ, constituting the putative structural homolog of Zα, the canonical Z-DNA/Z-RNA binding domain previously identified in the interferon-inducible RNA Adenosine-to-Inosine-editing deaminase, ADAR1p150. In contrast to Zα, which is separate from the protein segment responsible for catalysis, Zτ encompasses the active site tyrosine of topoII; a GTP-binding site and a GxxG sequence motif are in close proximity. Quantitative Zτ-Zα similarity comparisons and molecular docking with interaction scoring further supported the "B-Z-topoII hypothesis" and has led to an expanded mechanism for topoII function incorporating the recognition of Z-DNA segments ("Z-flipons") as an inherent and essential element. We further propose that the two Zτ domains of the topoII homodimer exhibit a single-turnover "conformase" activity on given G(ate) B-DNA segments ("Z-flipins"), inducing their transition to the left-handed Z-conformation. Inasmuch as the topoII-Z-DNA complexes are isomerase inactive, we infer that they fulfill important structural roles in key processes such as mitosis. Topoisomerases are preeminent targets of anti-cancer drug discovery, and we anticipate that detailed elucidation of their structural-functional interactions with Z-DNA and GTP will facilitate the design of novel, more potent and selective anti-cancer chemotherapeutic agents.
基因组 DNA 上的动态过程,如基因表达和细胞分裂,不可避免地导致拓扑学上的挑战,表现为缠绕、连环、结、“泡”、R 环和超螺旋及螺旋破坏的其他结果。有毒拓扑结构压力的解决是归因于 DNA 拓扑异构酶的功能。一个突出的例子是负超螺旋(nsc)尾随过程性酶,如 DNA 和 RNA 聚合酶。nscDNA 通过螺旋扭曲和缠绕的重新分布可以采用多种平衡状态,包括称为 Z-DNA 的左手双螺旋构象。三十年前,我们的一个实验室从细胞和胚胎中分离出一种蛋白质,它对 Z-DNA 的亲和力比 B-DNA 高 100 倍,并将其鉴定为拓扑异构酶 II(基因 Top2,与人类 UniProt 蛋白 TOP2A 和 TOP2B 同源)。GTP 进一步提高了与 Z-DNA 的亲和力和选择性,同时也抑制了异构酶的酶活性。提出了一种变构机制,其中拓扑异构酶 II 作为 Z-DNA 结合蛋白(ZBP)稳定拓扑(亚)域和相关多蛋白复合物的特定状态。我们现在通过对代表整个生命树的生物体的拓扑异构酶 II 可用蛋白质序列进行全面的生物信息学分析,探索了这种可能性。对这些序列的多重比对显示出极高的进化保守性,包括一个翼状螺旋蛋白片段,这里称为 Zτ,构成先前在干扰素诱导的 RNA 腺苷酸到肌苷酸编辑脱氨酶 ADAR1p150 中鉴定的典型 Z-DNA/Z-RNA 结合域 Zα 的假定结构同源物。与 Zα 不同,Zα 与负责催化的蛋白质片段分开,Zτ包含拓扑异构酶 II 的活性位点酪氨酸;一个 GTP 结合位点和一个 GxxG 序列基序非常接近。定量 Zτ-Zα 相似性比较和分子对接以及相互作用评分进一步支持了“B-Z-topoII 假说”,并导致了拓扑异构酶功能的扩展机制,将识别 Z-DNA 片段(“Z-翻转”)作为固有和基本元素纳入其中。我们进一步提出,拓扑异构酶 II 同源二聚体的两个 Zτ 结构域对特定的 G(ate)B-DNA 片段(“Z-翻转”)表现出单轮“构象酶”活性,诱导它们向左手 Z 构象转变。由于拓扑异构酶 II-Z-DNA 复合物是无异构酶活性的,我们推断它们在有丝分裂等关键过程中发挥重要的结构作用。拓扑异构酶是抗癌药物发现的主要靶点,我们预计详细阐明它们与 Z-DNA 和 GTP 的结构功能相互作用将有助于设计新型、更有效和选择性的抗癌化疗药物。