Department of Chemistry and Biochemistry, University of Nevada, Las Vegas, Las Vegas, United States.
Elife. 2024 Feb 12;13:e86168. doi: 10.7554/eLife.86168.
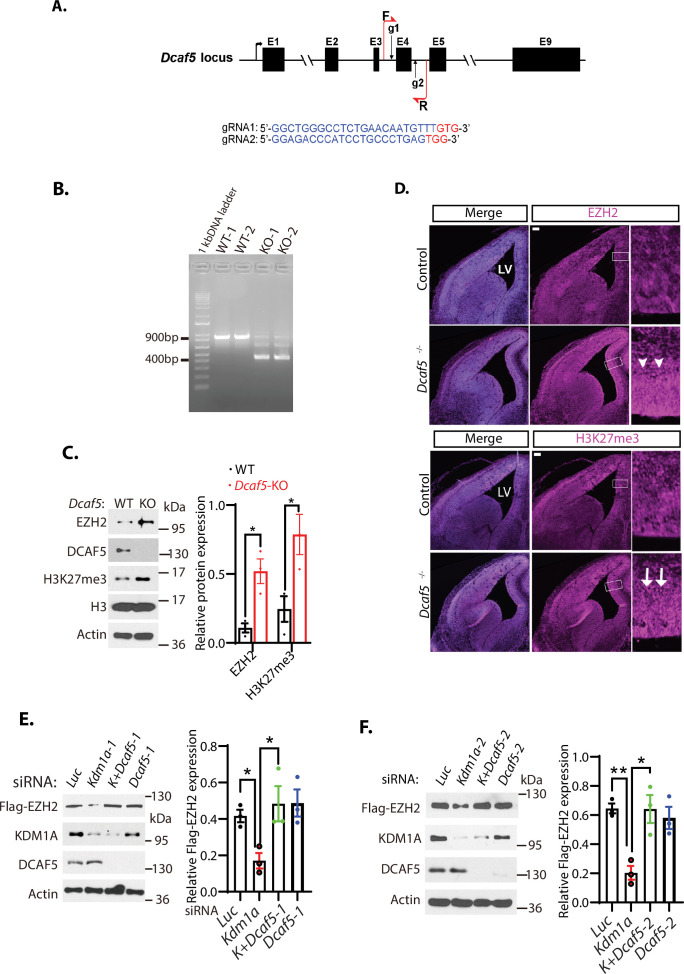
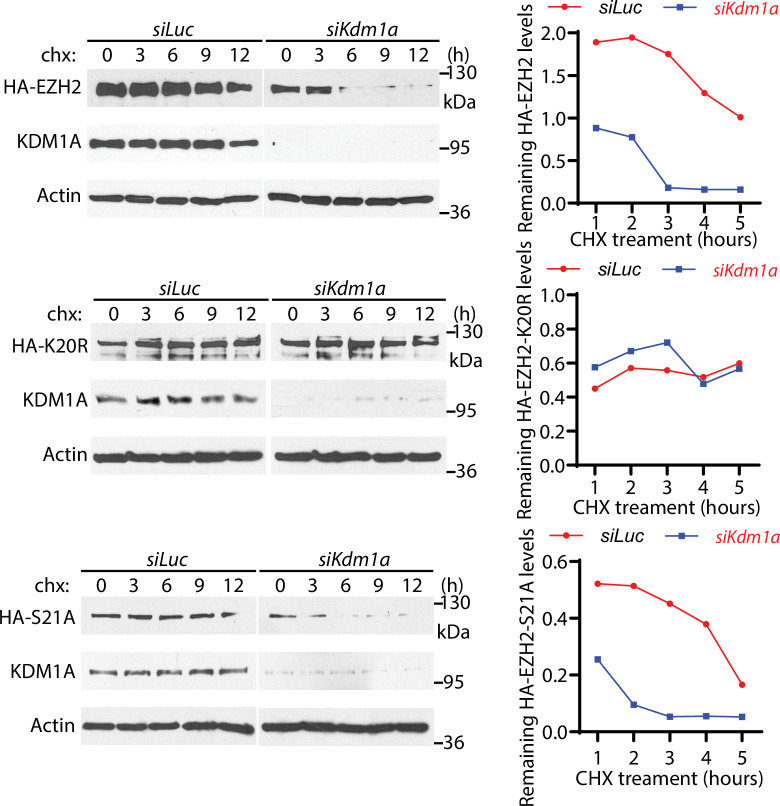
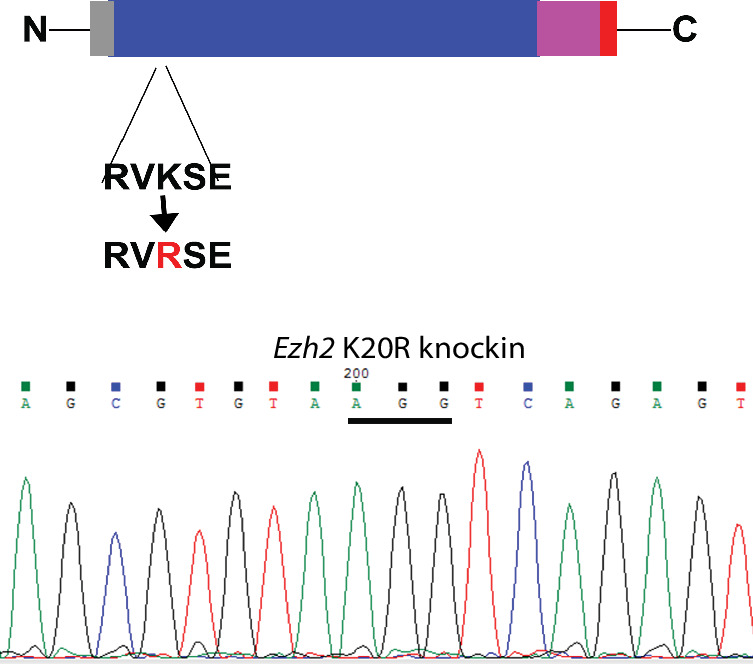
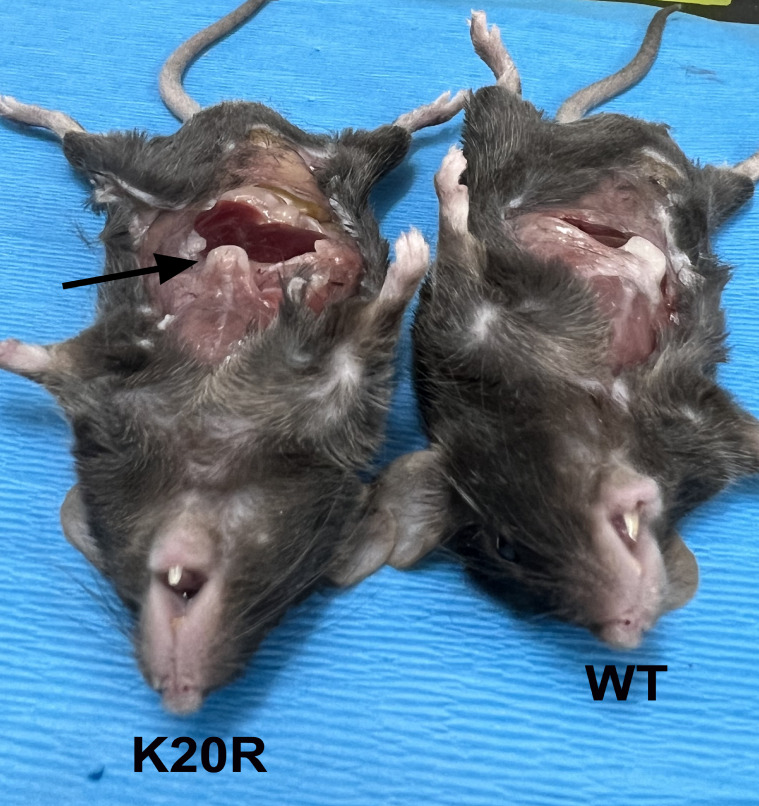
The Polycomb Repressive Complex 2 (PRC2) methylates H3K27 to regulate development and cell fate by transcriptional silencing. Alteration of PRC2 is associated with various cancers. Here, we show that mouse deletion causes a dramatic reduction of PRC2 proteins, whereas mouse null mutation of or results in PRC2 accumulation and increased H3K27 trimethylation. The catalytic subunit of PRC2, EZH2, is methylated at lysine 20 (K20), promoting EZH2 proteolysis by L3MBTL3 and the CLR4 ubiquitin ligase. KDM1A (LSD1) demethylates the methylated K20 to stabilize EZH2. K20 methylation is inhibited by AKT-mediated phosphorylation of serine 21 in EZH2. Mouse mutants develop hepatosplenomegaly associated with high GFI1B expression, and mutant bone marrows expand hematopoietic stem cells and downstream hematopoietic populations. Our studies reveal that EZH2 is regulated by methylation-dependent proteolysis, which is negatively controlled by AKT-mediated S21 phosphorylation to establish a methylation-phosphorylation switch to regulate the PRC2 activity and hematopoiesis.
多梳抑制复合物 2(PRC2)通过转录沉默将 H3K27 甲基化,从而调节发育和细胞命运。PRC2 的改变与各种癌症有关。在这里,我们表明,小鼠 缺失导致 PRC2 蛋白的显著减少,而 或 缺失导致 PRC2 积累和 H3K27 三甲基化增加。PRC2 的催化亚基 EZH2 在赖氨酸 20(K20)处被甲基化,促进 L3MBTL3 和 CLR4 泛素连接酶对 EZH2 的蛋白水解。KDM1A(LSD1)通过去甲基化将甲基化的 K20 转化为稳定的 EZH2。EZH2 的 K20 甲基化被 AKT 介导的丝氨酸 21 磷酸化抑制。小鼠 突变体表现出肝脾肿大,伴有高表达的 GFI1B,而 突变体骨髓扩增造血干细胞和下游造血群体。我们的研究表明,EZH2 受依赖于甲基化的蛋白水解调节,该蛋白水解受 AKT 介导的 S21 磷酸化负调控,以建立一个甲基化-磷酸化开关来调节 PRC2 活性和造血。