Lin Che-Yi, Marlétaz Ferdinand, Pérez-Posada Alberto, Martínez-García Pedro Manuel, Schloissnig Siegfried, Peluso Paul, Conception Greg T, Bump Paul, Chen Yi-Chih, Chou Cindy, Lin Ching-Yi, Fan Tzu-Pei, Tsai Chang-Tai, Gómez Skarmeta José Luis, Tena Juan J, Lowe Christopher J, Rank David R, Rokhsar Daniel S, Yu Jr-Kai, Su Yi-Hsien
Institute of Cellular and Organismic Biology, Academia Sinica, Taipei, Taiwan.
Center for Life's Origins and Evolution, Department of Genetics, Evolution and Environment, University College London, London, United Kingdom.
PLoS Biol. 2024 Jun 3;22(6):e3002661. doi: 10.1371/journal.pbio.3002661. eCollection 2024 Jun.
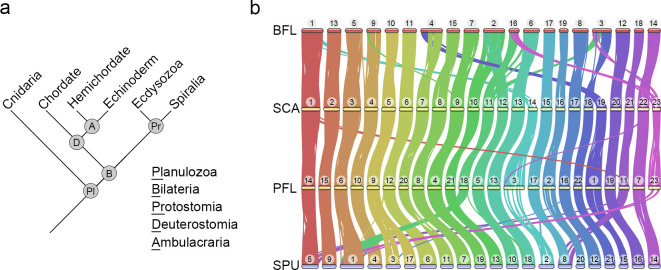
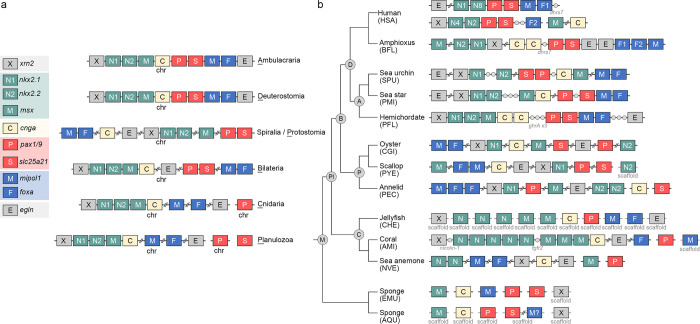
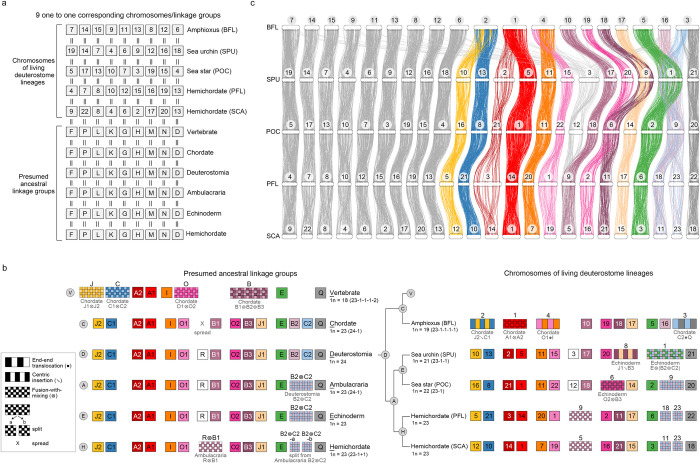
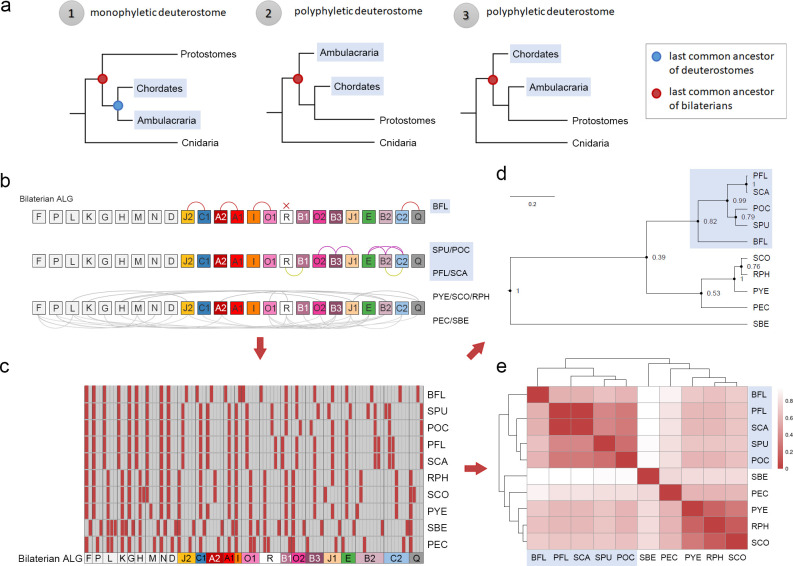
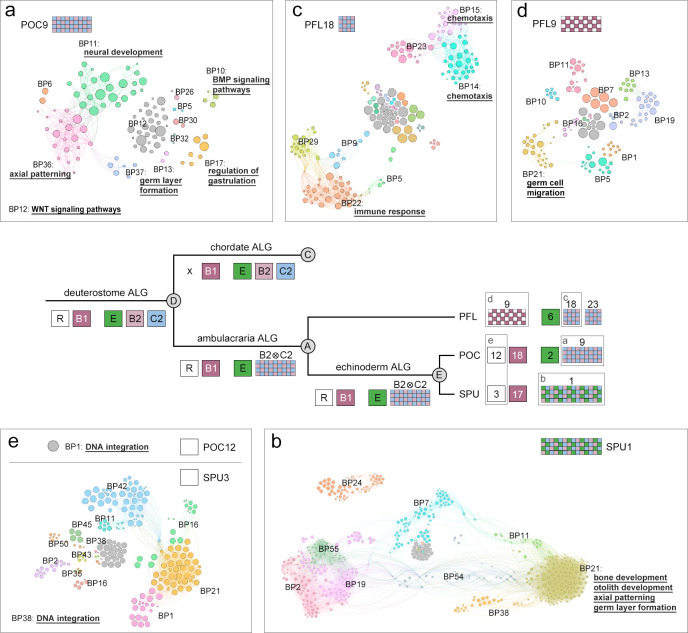
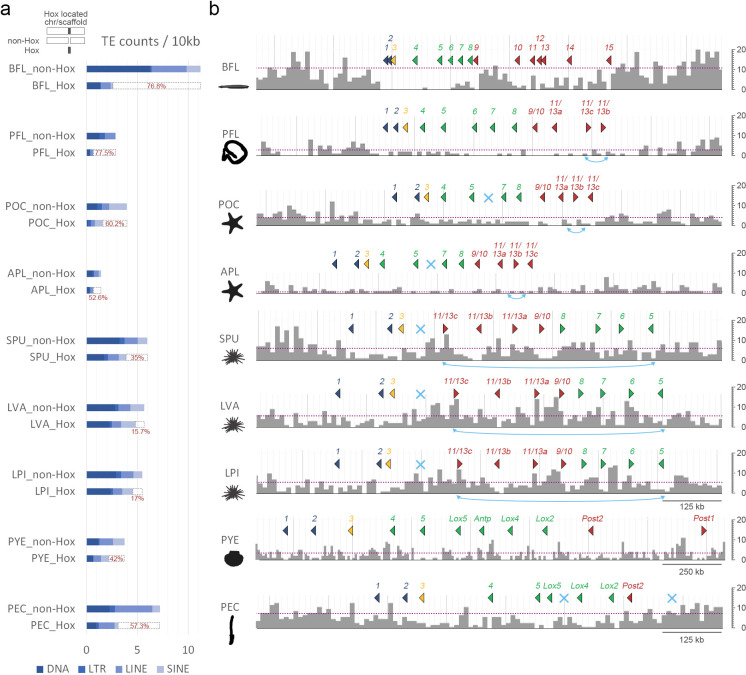
Deuterostomes are a monophyletic group of animals that includes Hemichordata, Echinodermata (together called Ambulacraria), and Chordata. The diversity of deuterostome body plans has made it challenging to reconstruct their ancestral condition and to decipher the genetic changes that drove the diversification of deuterostome lineages. Here, we generate chromosome-level genome assemblies of 2 hemichordate species, Ptychodera flava and Schizocardium californicum, and use comparative genomic approaches to infer the chromosomal architecture of the deuterostome common ancestor and delineate lineage-specific chromosomal modifications. We show that hemichordate chromosomes (1N = 23) exhibit remarkable chromosome-scale macrosynteny when compared to other deuterostomes and can be derived from 24 deuterostome ancestral linkage groups (ALGs). These deuterostome ALGs in turn match previously inferred bilaterian ALGs, consistent with a relatively short transition from the last common bilaterian ancestor to the origin of deuterostomes. Based on this deuterostome ALG complement, we deduced chromosomal rearrangement events that occurred in different lineages. For example, a fusion-with-mixing event produced an Ambulacraria-specific ALG that subsequently split into 2 chromosomes in extant hemichordates, while this homologous ALG further fused with another chromosome in sea urchins. Orthologous genes distributed in these rearranged chromosomes are enriched for functions in various developmental processes. We found that the deeply conserved Hox clusters are located in highly rearranged chromosomes and that maintenance of the clusters are likely due to lower densities of transposable elements within the clusters. We also provide evidence that the deuterostome-specific pharyngeal gene cluster was established via the combination of 3 pre-assembled microsyntenic blocks. We suggest that since chromosomal rearrangement events and formation of new gene clusters may change the regulatory controls of developmental genes, these events may have contributed to the evolution of diverse body plans among deuterostomes.
后口动物是一类单系群动物,包括半索动物门、棘皮动物门(合称为有腕动物)和脊索动物门。后口动物身体结构的多样性使得重建它们的祖先状态以及解读驱动后口动物谱系多样化的基因变化变得具有挑战性。在这里,我们生成了两种半索动物物种——黄殖翼柱头虫和加州裂体柱头虫的染色体水平基因组组装,并使用比较基因组方法来推断后口动物共同祖先的染色体结构,并描绘谱系特异性的染色体修饰。我们表明,与其他后口动物相比,半索动物染色体(1N = 23)表现出显著的染色体规模的宏观共线性,并且可以源自24个后口动物祖先连锁群(ALG)。这些后口动物ALG又与先前推断的两侧对称动物ALG相匹配,这与从最后一个共同的两侧对称动物祖先到后口动物起源的相对较短的过渡相一致。基于这个后口动物ALG互补体,我们推断了在不同谱系中发生的染色体重排事件。例如,一个融合与混合事件产生了一个有腕动物特异性的ALG,该ALG随后在现存的半索动物中分裂成两条染色体,而这个同源的ALG在海胆中进一步与另一条染色体融合。分布在这些重排染色体中的直系同源基因在各种发育过程中的功能上富集。我们发现深度保守的Hox簇位于高度重排的染色体中,并且这些簇的维持可能是由于簇内转座元件的密度较低。我们还提供证据表明,后口动物特异性的咽基因簇是通过3个预先组装的微共线性模块的组合建立的。我们认为由于染色体重排事件和新基因簇的形成可能会改变发育基因的调控控制,这些事件可能促成了后口动物中多样化身体结构的进化。