Department of Molecular Physiology and Biological Physics, Center for Membrane and Cell Physiology, University of Virginia, Charlottesville, United States.
Department of Chemistry, The University of Tennessee, Knoxville, United States.
Elife. 2024 Jun 5;12:RP89306. doi: 10.7554/eLife.89306.
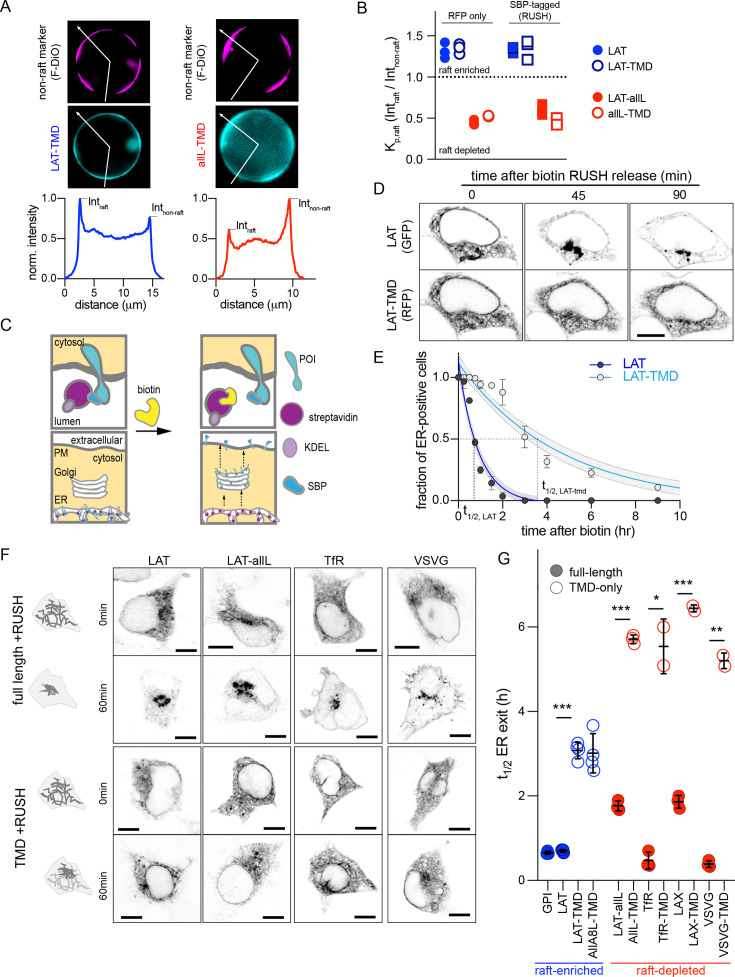
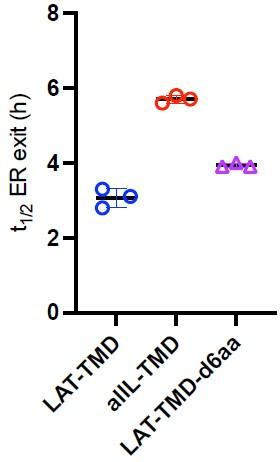
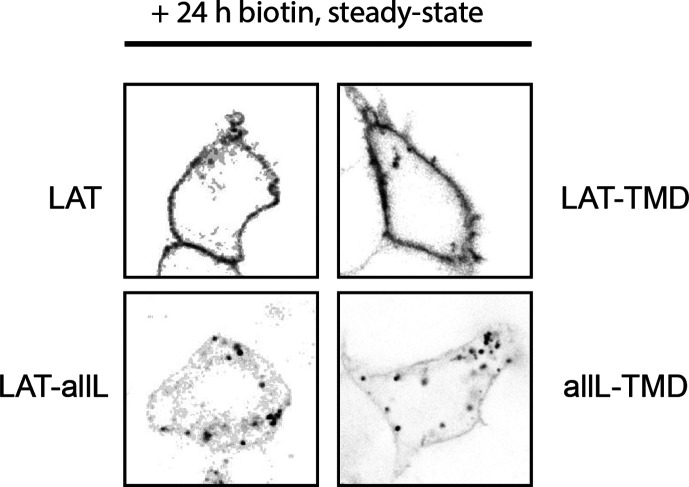
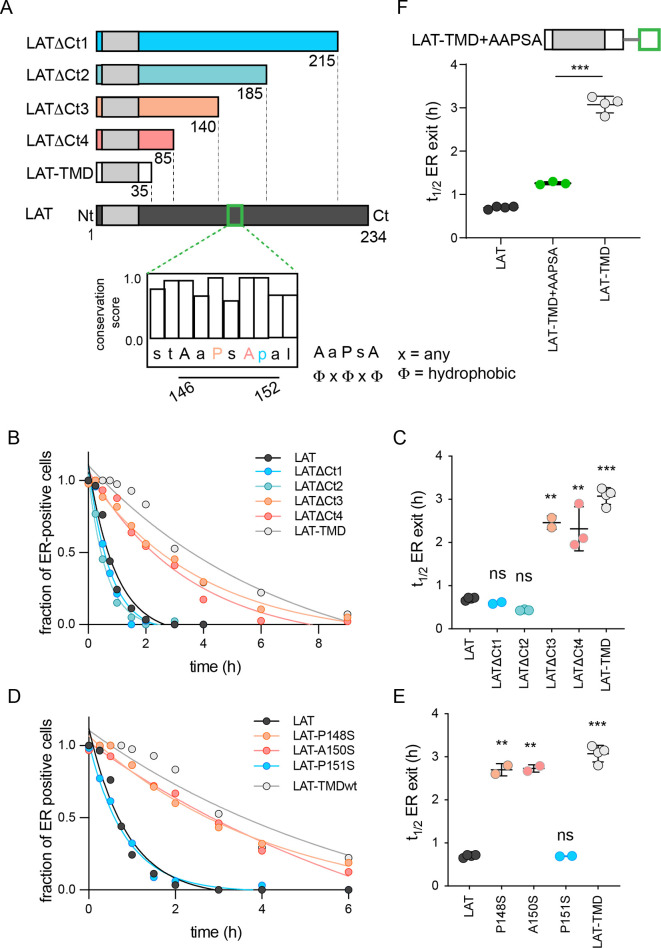
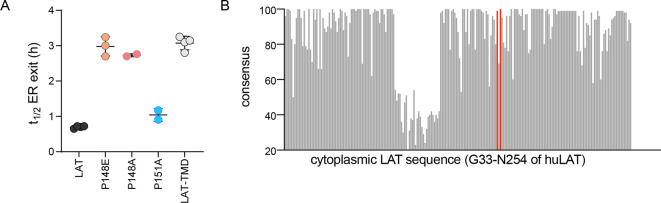
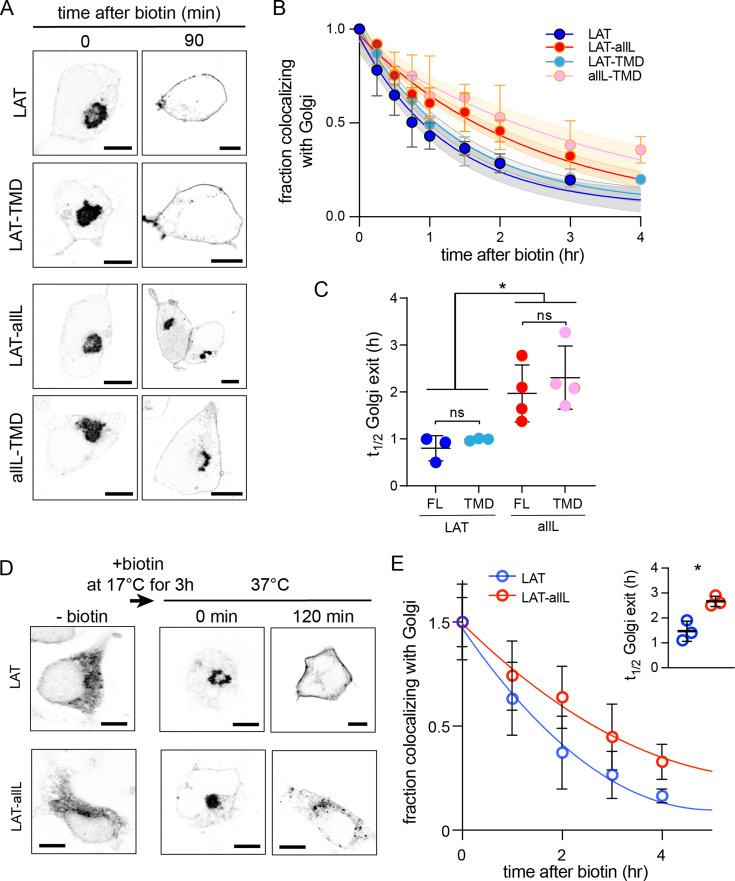
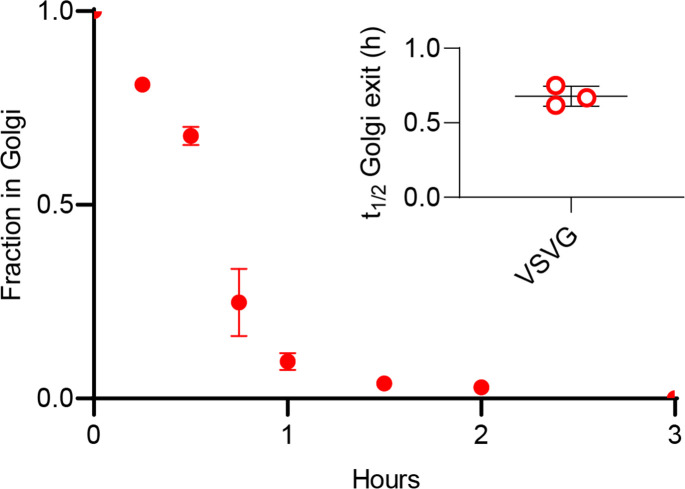
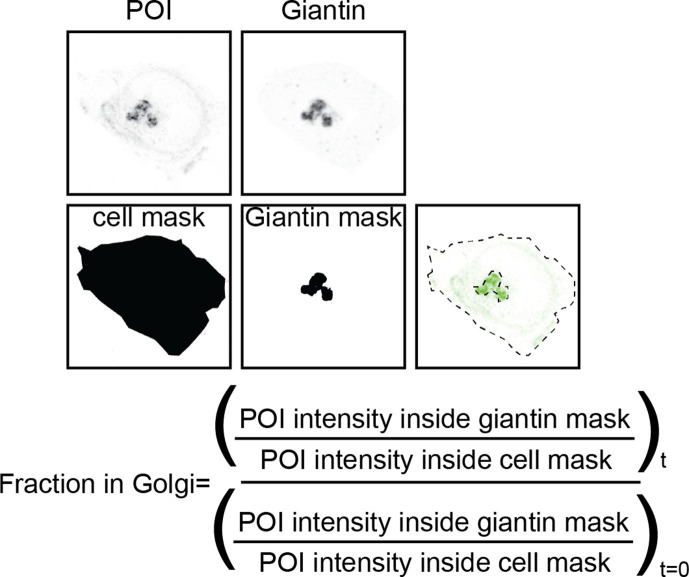
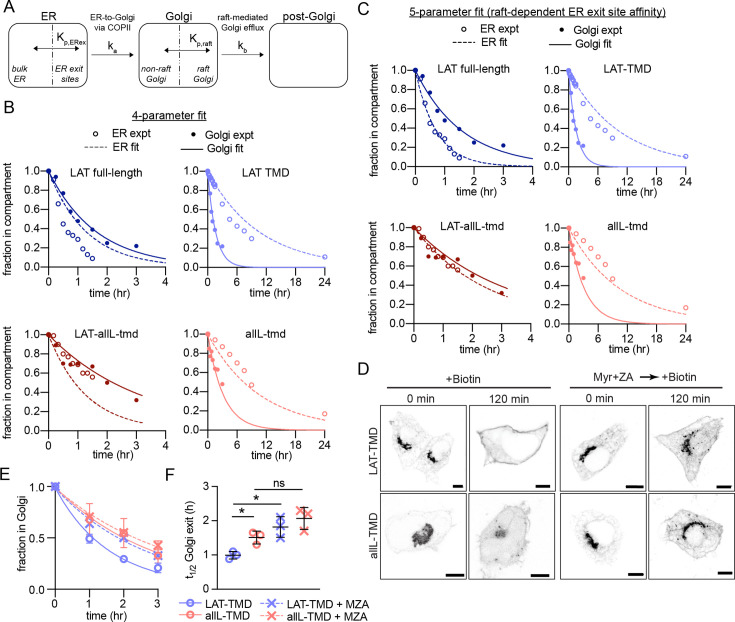
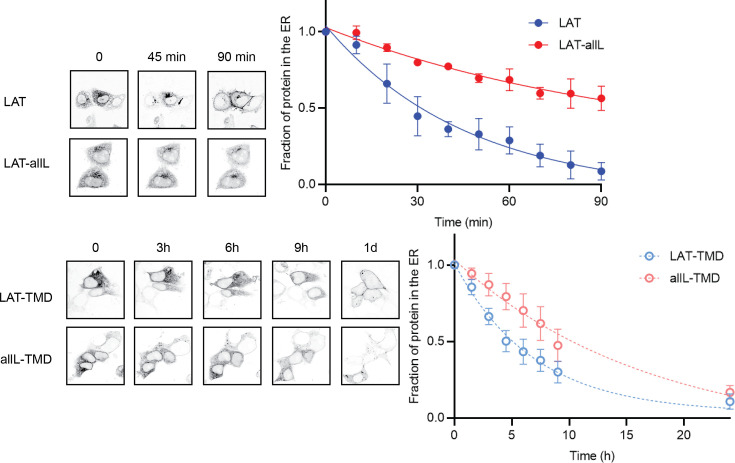
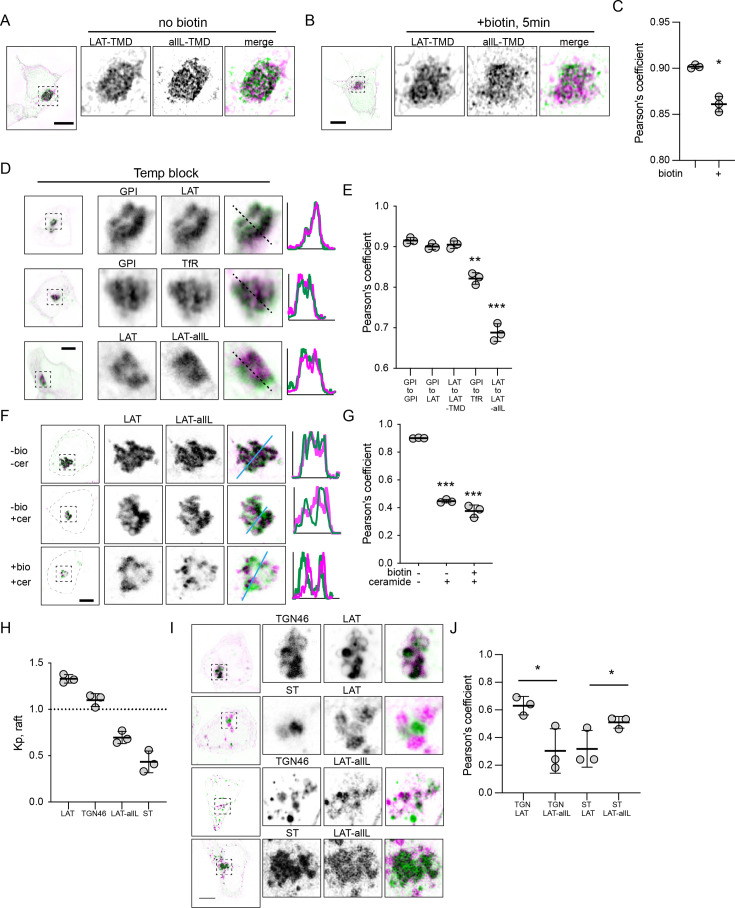
The organelles of eukaryotic cells maintain distinct protein and lipid compositions required for their specific functions. The mechanisms by which many of these components are sorted to their specific locations remain unknown. While some motifs mediating subcellular protein localization have been identified, many membrane proteins and most membrane lipids lack known sorting determinants. A putative mechanism for sorting of membrane components is based on membrane domains known as lipid rafts, which are laterally segregated nanoscopic assemblies of specific lipids and proteins. To assess the role of such domains in the secretory pathway, we applied a robust tool for synchronized secretory protein traffic (RUSH, etention sing elective ooks) to protein constructs with defined affinity for raft phases. These constructs consist solely of single-pass transmembrane domains (TMDs) and, lacking other sorting determinants, constitute probes for membrane domain-mediated trafficking. We find that while raft affinity can be sufficient for steady-state PM localization, it is not sufficient for rapid exit from the endoplasmic reticulum (ER), which is instead mediated by a short cytosolic peptide motif. In contrast, we find that Golgi exit kinetics are highly dependent on raft affinity, with raft preferring probes exiting the Golgi ~2.5-fold faster than probes with minimal raft affinity. We rationalize these observations with a kinetic model of secretory trafficking, wherein Golgi export can be facilitated by protein association with raft domains. These observations support a role for raft-like membrane domains in the secretory pathway and establish an experimental paradigm for dissecting its underlying machinery.
真核细胞的细胞器维持着其特定功能所需的独特蛋白质和脂质组成。许多这些成分被分拣到其特定位置的机制仍然未知。虽然已经确定了一些介导亚细胞蛋白质定位的基序,但许多膜蛋白和大多数膜脂质缺乏已知的分拣决定因素。一种用于分拣膜成分的假定机制基于称为脂筏的膜域,它们是特定脂质和蛋白质侧向分隔的纳米级组装体。为了评估这些域在分泌途径中的作用,我们应用了一种用于同步分泌蛋白运输的强大工具(RUSH,保留选择书籍)来研究具有确定的筏相亲和力的蛋白质构建体。这些构建体仅由单通道跨膜结构域 (TMD) 组成,并且由于缺乏其他分拣决定因素,它们构成了用于膜域介导的运输的探针。我们发现,虽然筏亲和力足以维持 PM 的稳定定位,但不足以快速从内质网 (ER) 中逸出,而这是通过短的细胞质肽基序介导的。相比之下,我们发现高尔基体出口动力学高度依赖于筏亲和力,与最小筏亲和力的探针相比,具有筏亲和力的探针从高尔基体中逸出的速度快 2.5 倍。我们用分泌运输的动力学模型来解释这些观察结果,其中高尔基体出口可以通过蛋白质与筏域的结合来促进。这些观察结果支持了类似筏的膜域在分泌途径中的作用,并建立了用于剖析其潜在机制的实验范例。