Department of Molecular, Cell and Developmental Biology, University of California Los Angeles, Los Angeles, CA, USA.
Eli and Edythe Broad Center for Regenerative Medicine and Stem Cell Research, University of California Los Angeles, Los Angeles, CA, USA.
Nature. 2024 Jun;630(8016):412-420. doi: 10.1038/s41586-024-07478-x. Epub 2024 Jun 5.
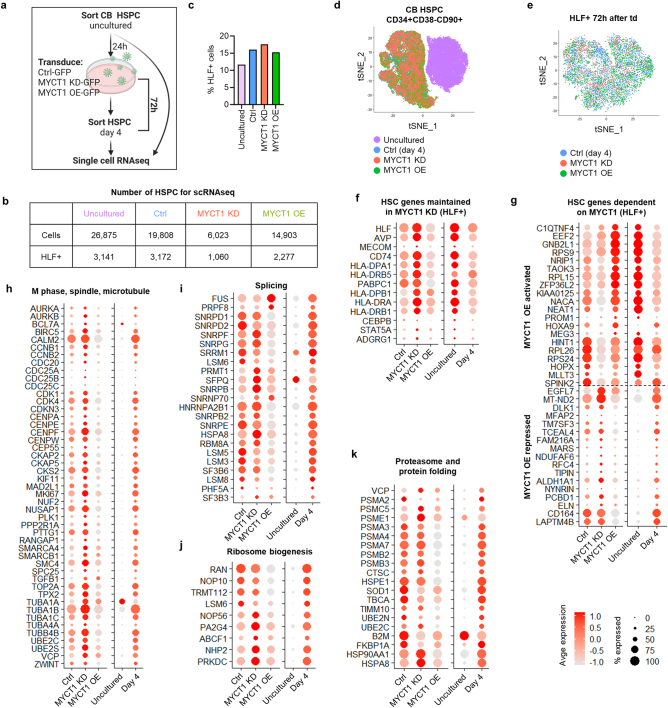
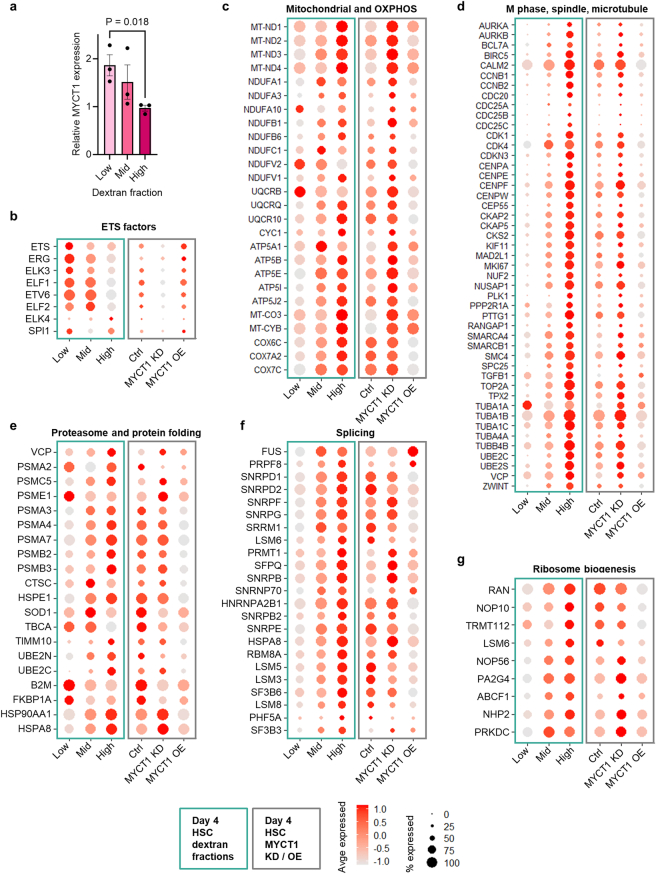
The processes that govern human haematopoietic stem cell (HSC) self-renewal and engraftment are poorly understood and challenging to recapitulate in culture to reliably expand functional HSCs. Here we identify MYC target 1 (MYCT1; also known as MTLC) as a crucial human HSC regulator that moderates endocytosis and environmental sensing in HSCs. MYCT1 is selectively expressed in undifferentiated human haematopoietic stem and progenitor cells (HSPCs) and endothelial cells but becomes markedly downregulated during HSC culture. Lentivirus-mediated knockdown of MYCT1 prevented human fetal liver and cord blood (CB) HSPC expansion and engraftment. By contrast, restoring MYCT1 expression improved the expansion and engraftment of cultured CB HSPCs. Single-cell RNA sequencing of human CB HSPCs in which MYCT1 was knocked down or overexpressed revealed that MYCT1 governs important regulatory programmes and cellular properties essential for HSC stemness, such as ETS factor expression and low mitochondrial activity. MYCT1 is localized in the endosomal membrane in HSPCs and interacts with vesicle trafficking regulators and signalling machinery. MYCT1 loss in HSPCs led to excessive endocytosis and hyperactive signalling responses, whereas restoring MYCT1 expression balanced culture-induced endocytosis and dysregulated signalling. Moreover, sorting cultured CB HSPCs on the basis of lowest endocytosis rate identified HSPCs with preserved MYCT1 expression and MYCT1-regulated HSC stemness programmes. Our work identifies MYCT1-moderated endocytosis and environmental sensing as essential regulatory mechanisms required to preserve human HSC stemness. Our data also pinpoint silencing of MYCT1 as a cell-culture-induced vulnerability that compromises human HSC expansion.
调控人类造血干/祖细胞(HSC)自我更新和植入的过程尚未被充分了解,并且难以在培养物中重现,以可靠地扩增功能性 HSC。在这里,我们鉴定出 MYC 靶蛋白 1(MYCT1;也称为 MTLC)是一种关键的人类 HSC 调节因子,可调节 HSC 中的内吞作用和环境感应。MYCT1 在未分化的人类造血干/祖细胞(HSPC)和内皮细胞中选择性表达,但在 HSC 培养过程中明显下调。慢病毒介导的 MYCT1 敲低可阻止人胎肝和脐血(CB)HSPC 的扩增和植入。相比之下,恢复 MYCT1 表达可改善培养的 CB HSPC 的扩增和植入。敲低或过表达 MYCT1 的人 CB HSPC 的单细胞 RNA 测序显示,MYCT1 调控重要的调控程序和对 HSC 干性至关重要的细胞特性,例如 ETS 因子表达和低线粒体活性。MYCT1 在 HSPC 中定位于内体膜,并与囊泡运输调节剂和信号机制相互作用。HSPC 中的 MYCT1 缺失导致过度的内吞作用和过度活跃的信号反应,而恢复 MYCT1 表达则平衡了培养诱导的内吞作用和失调的信号反应。此外,基于最低内吞作用率对培养的 CB HSPC 进行分选,鉴定出具有保留的 MYCT1 表达和 MYCT1 调节的 HSC 干性程序的 HSPC。我们的工作确定了 MYCT1 调节的内吞作用和环境感应作为维持人类 HSC 干性所必需的重要调节机制。我们的数据还指出,MYCT1 的沉默是一种细胞培养诱导的脆弱性,会损害人类 HSC 的扩增。