Division of Radiation and Cancer Biology, Department of Radiation Oncology, Stanford University School of Medicine, Stanford, CA 94305, USA.
Sci Adv. 2024 Aug 2;10(31):eadn4682. doi: 10.1126/sciadv.adn4682. Epub 2024 Jul 31.
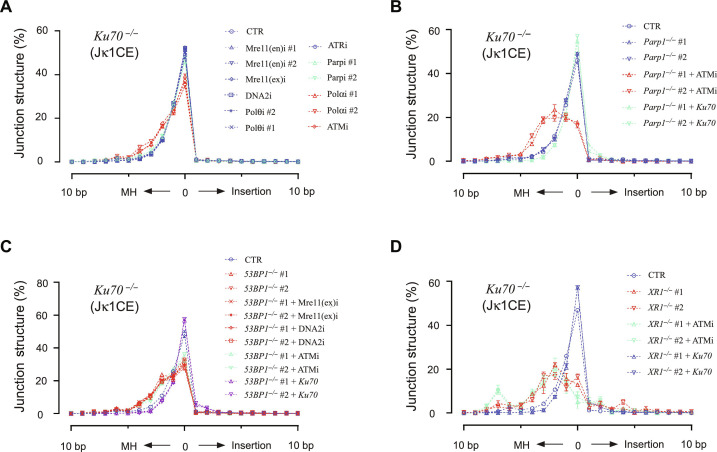
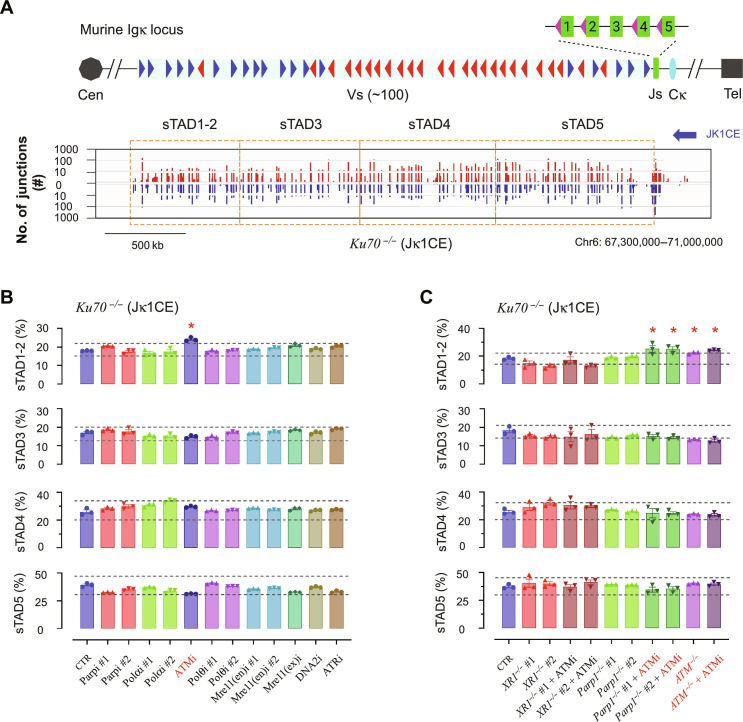
G-G phase alternative end joining (A-EJ) is a recently defined mutagenic pathway characterized by resected deletion and translocation joints that are predominantly direct and are distinguished from A-EJ in cycling cells that rely much more on microhomology-mediated end joining (MMEJ). Using chemical and genetic approaches, we systematically evaluate potential A-EJ factors and DNA damage response (DDR) genes to support this mechanism by mapping the repair fates of RAG1/2-initiated double-strand breaks in the context of Igκ locus V-J recombination and chromosome translocation. Our findings highlight a polymerase theta-independent Parp1-XRCC1/LigIII axis as central A-EJ components, supported by 53BP1 in the context of an Ataxia-telangiectasia mutated (ATM)-activated DDR. Mechanistically, we demonstrate varied changes in short-range resection, MMEJ, and translocation, imposed by compromising specific DDR activities, which include polymerase alpha, Ataxia-telangiectasia and Rad3-related (ATR), DNA2, and Mre11. This study advances our understanding of DNA damage repair within the 53BP1 regulatory domain and the RAG1/2 postcleavage complex.
G1-G2 期交替末端连接(A-EJ)是一种新定义的诱变途径,其特征是切除和易位连接,主要是直接的,与循环细胞中的 A-EJ 不同,后者更依赖于微同源介导的末端连接(MMEJ)。我们使用化学和遗传方法,通过在免疫球蛋白κ基因座 V-J 重组和染色体易位的背景下绘制 RAG1/2 起始的双链断裂的修复命运,系统地评估潜在的 A-EJ 因子和 DNA 损伤反应(DDR)基因,以支持这种机制。我们的发现强调了一个聚合酶θ独立的 Parp1-XRCC1/LigIII 轴作为中央 A-EJ 成分,在 ATM 激活的 DDR 背景下由 53BP1 支持。从机制上讲,我们通过削弱特定的 DDR 活性,包括聚合酶α、共济失调毛细血管扩张症和 Rad3 相关(ATR)、DNA2 和 Mre11,证明了短距离切除、MMEJ 和易位的变化。这项研究推进了我们对 53BP1 调节域和 RAG1/2 后切割复合物内 DNA 损伤修复的理解。